Spekroflormetrik metotla insan serum albumin proteini ile neohesperidin dihidrokalkon ve maprotilin hidroklorür arasındaki etkileşimlerin incelenmesi
Tam metin
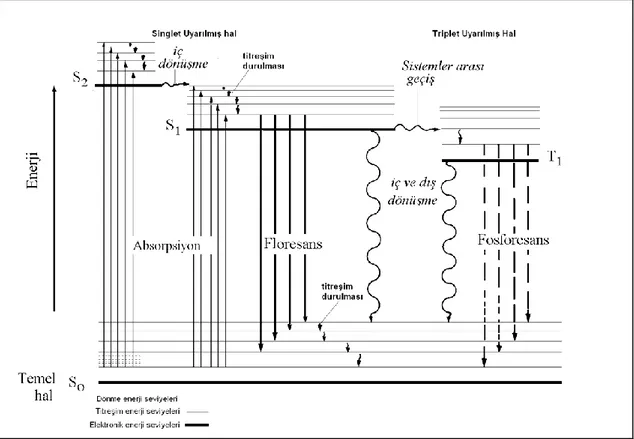
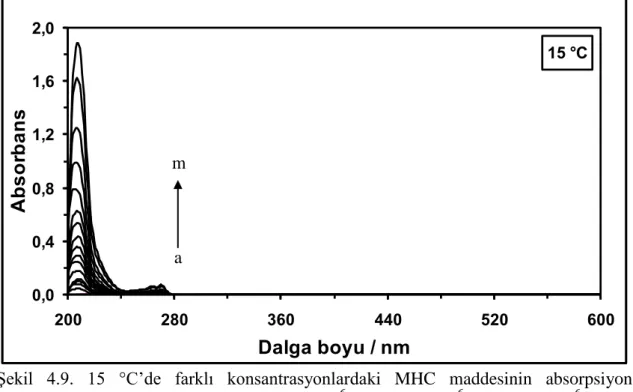
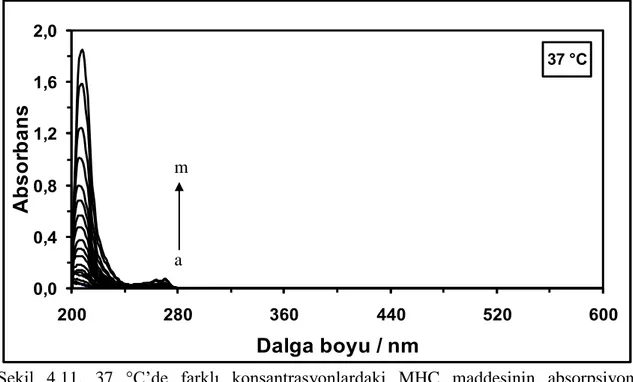
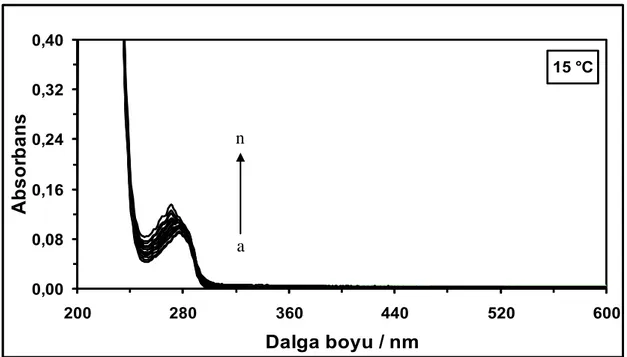
Şekil






Benzer Belgeler
[Androcur ] - [安得卡錠] 返回 藥品介紹 藥師 藥劑部藥師 發佈日期 2010/02/11 <藥物效用>
Azmun, Dede Korkut Kitabı’nın üçüncü yazmasının içinde iki yeni boy olduğunu düşünürken (1. Salur Kazan’ın Aras Suyu ile Kars kal’asını aldığı boy
Teorik İlaç Etken Madde Tasarımı Sentezi YENİ İLAÇ ETKEN MADDESİ Biyolojik Etkinin Gözlenmesi *** Esin AKI BİLGİSAYAR DESTEKLİ İLAÇ TASARIMI
otre Dame de Sion Lisesi, 150 yıldan beri rahibeleri, Fransızca öğretmesi, 10 yıl öncesine kadar sadece kız öğrenci alırken bir devrim yapıp erkek öğrencilere de
Rinaldo Marmara, Ahmet Hamdi Tanpınar’ın geçen yüzyılda “şehirlerin bahçeleri” olduğunu söylediği mezarlıklar arasından Pera sosyetesinin buluşma yeri olan,
Ele alınan onaltı örnek arasında dörtgen, yassı, dairesel gibi Ortaçağ döneminin en sık karşılaşılan tipleri, üç bıçaklı (kanatlı) diye adlandırılan
Elde edilen bulgulara göre öğretmenler, eğitim sistemindeki kademe yapısının 4+4+4 şeklinde değil 1998 yılı öncesindeki uygulamayı anımsatan bir biçimde
Ortalamalarının Hangi Gruplar Arasında Farklılık Gösterdiğini Belirleyebilmek İçin Yapılan LSD Testi Sonuçları ...67 Tablo 20: Bireyler Arası İlişkiler
