T.C.
SELÇUK ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ
MASTĠTĠSLĠ ĠNEKLERDEN ĠZOLE EDĠLEN
STAPHYLOCOCCUS AUREUS SUġLARININ
BAZI VĠRULANS FAKTÖRLERE [SPA (Xr ve IgG BAĞLANMA BÖLGESĠ), COA,
clfA] GÖRE TĠPLENDĠRĠLMESĠ
Rukiye AYDINALP YÜKSEK LĠSANS TEZĠ
Biyoloji Anabilim Dalı
Mayıs-2015 KONYA Her Hakkı Saklıdır
iv
ÖZET YÜKSEK LĠSANS
MASTĠTĠSLĠ ĠNEKLERDEN ĠZOLE EDĠLEN STAPHYLOCOCCUS AUREUS SUġLARININ BAZI VĠRULANS FAKTÖRLERE [SPA (Xr ve IgG BAĞLANMA
BÖLGESĠ), COA, clfA] GÖRE TĠPLENDĠRĠLMESĠ Rukiye AYDINALP
Selçuk Üniversitesi Fen Bilimleri Enstitüsü Biyoloji Anabilim Dalı
DanıĢman: Doç. Dr. Emine ARSLAN 2015, 41
Jüri
Doç. Dr. Emine ARSLAN Doç. Dr. Rüstem DUMAN Yrd. Doç. Dr. Hilal ARIKOĞLU
Mastitis sığırlarda çoğunlukla Staphylococcus aureus’un neden olduğu yaygın hastalıklardan biridir. S. aureus sığırlarda mastitise neden olan virulans faktörler olarak rol oynayan birçok protein kodlar. Bu çalıĢmada mastitisli ineklerden izole edilen 98 S. aureus suĢunda PCR ile spa-X, spa-IgG, clfA ve coa gibi virulans genlerin varlığı/yokluğu belirlenmiĢ ve gözlemlenen genlerin büyüklüklerinin analizi yapılmıĢtır. PCR sonuçlarına göre 98 örneğin %98.97’i spa-X genini, %80.61’i spa-IgG genini, %76.53’ü clfA genini, %74.48’i coa genini bulundurduğu sonucu elde edilmiĢtir. Ayrıca spa-X geninde 100 bç, 250 bç, 300 bç, spa-IgG geninde 800 bç ve 900 bç clfA geninde 1000 bç coa geninde 600 bç, 700 bç ve 850 bç büyüklüğünde bantlar elde edilmiĢtir. Sonuç olarak mastitisli ineklerden izole edilen S. aureus suĢlarında bu virulans genlerinin prevelansının yüksek olduğu belirlenmiĢtir. Bu virulans genlerin tiplendirilmesi gelecekteki aĢı çalıĢmalarına ıĢık tutabilir.
v
ABSTRACT MS THESIS
TYPĠNG OF STAPHYLOCOCCUS AUREUS STRAĠNS ĠSOLATED FROM BOVĠNE MASTĠTĠS ACCORDĠNG TO SOME VĠRULANCE FACTORS
[SPA (Xr and IgG BĠNDĠNG REGĠON), COA, clfA] Rukiye AYDINALP
The Graduate School of Natural And Applied Science of Selçuk Unıversity Biology Branch
Advisor: Doç. Dr. Emine ARSLAN 2015, 41
Jury
Assoc. Prof. Dr. Emine ARSLAN Assoc. Prof. Dr. Rüstem DUMAN Asst. Prof. Dr. Hilal ARIKOĞLU
Mastitis is one of the common diseases caused mostly Staphylococcus aureus in dairy cows. S. aureus encodes many proteins that act as virulence factors leading to mastitis in cattle. In this study, existence/absence of virulans genes like spa-X, spa-IgG, clfA and coa have been determined in 98 S. aureus strain isolated from bovine mastitis and the quantity of these genes has been analysed by PCR. According to the results of PCR, 98 examples consist 98.97% spa-X gene, 80.61% spa-lgG gene, 76.53% clfA gene 74.48% coa gene. In addition, 100 bç, 250 bç, 300 bç have been found in spa-X gene; 800 bç and 900 bç have been found in spa-lgG gene; 1000 bç have been found in clfA gene; 600 bç, 700 bç and 850 bç have been found in coa gene. Consequently, in S. aureus strains isolated from bovine mastitis, the prevalence of these virulence genes were determined to be high. Tayping of this virulence genes may shed light/on future vaccine studies.
vi
ÖNSÖZ
Tez çalışmamın her aşamasında bilgi ve önerilerini esirgemeyen, her konuda bana yol gösteren, danışman hocam Sayın Doç. Dr. Emine ARSLAN’a sonsuz teşekkürlerimi sunarım. Deney aşamalarında bilgi ve tecrübelerini benimle paylaşan yardımlarını esirgemeyen Elif GÜLBAHÇE MUTLU’ya, ayrıca bana bu süre boyunca her konuda destek olan, sıkıntılarımı paylaşan, maddi ve manevi desteklerini esirgemeyen sevgili aileme teşekkür ederim.
Ayrıca 12201038 nolu proje ile tezimin yürütülmesinde araç gereç ve maddi Olanak sağlayan S.Ü. BAP Koordinatörlüğü’ne teşekkürlerimi sunarım.
Rukiye AYDINALP KONYA-2015
vii ĠÇĠNDEKĠLER ÖZET ... iv ABSTRACT ... v ÖNSÖZ ... vi ĠÇĠNDEKĠLER ... vii
SĠMGELER VE KISALTMALAR ... viii
1. GĠRĠġ ... 1
2. KAYNAK ARAġTIRMASI ... 5
3. MATERYAL VE YÖNTEM ... 15
3.1. Materyal ... 15
3.1.1. Kimyasal maddeler ve besiyerleri ... 15
3.1.2. Tampon ve çözeltiler ... 15
3.2. Metod ... 16
3.2.1. Çalışmada kullanılan bakteri kültürleri ... 16
3.2.2. Bakteriden genomik DNA izolasyonu ... 16
3.2.3. PCR uygulaması ... 16
3.2.4. S. aureus suşlarının PCR ürünlerinin agaroz jel elektroforezi ... 17
3.2.5. Veri analizi ... 18
4. ARAġTIRMA SONUÇLARI VE TARTIġMA ... 19
4.1. S. aureus suşunda spa-X, spa-IgG, clfA ve coa genlerinin PCR sonuçları ... 19
5. SONUÇLAR VE ÖNERĠLER ... 33
KAYNAKLAR ... 34
viii
SĠMGELER VE KISALTMALAR
ark : Arkadaşları
bç : Baz çifti
ClfA : Klamping faktör-A ClfB : Klamping faktör-B CMT : California mastitis test
CNA : Kolajen bağlayıcı protein geni
Coa : Koagülaz
CRF : Coagulase-reacting factor
CTAB : Cetyl Trimethyl Ammonium Bromide dATP : Deoksi adenozin trifosfat
dCTP : Deoksi sitidin trifosfat dGTP : Deoksi guanozin trifosfat DNA : Deoksiribonükleik asit dNTP : dATP, dTTP, dCTP, dGTP dTTP : Deoksi timidin trifosfat EDTA : Etilendiamin tetraasetikasit EFP : Fibrinojen bağlayıcı protein geni FnbA : Fibronektin bağlayıcı-A
FnbB : Fibronektin bağlayıcı-B HCl : Hidroklorik asit
KNS : Koagülaz negatif stafilokok
Mg : Miligram
MgCl : Magnezyum klorür
ml : Mililitre
NB : Nutrient broth
ºC : Santgrad derece
PCR : Polimeraz zincir reaksiyonu
Ph : Power of hydrogen (Hidrojen gücü)
rpm : Revolutions per minute (Dakikadaki devir sayısı) rRNA : Ribozomal ribonükleik asit
S. aureus : Staphylococcus aureus
Scp : Staphopain
SDS : Sodyum dodesil sülfat
Spa-IgG : protein A İmmunoglobulin bağlanma bölgesi Spa : Stafilokokal protein A
Spa-X : Protein A X bölgesi SspA : Stafilokokal serin proteaz SspP : Sistein proteaz
TAE : Tris-asetik asit-EDTA
TE : Tris-EDTA
Tm : Erime noktası
TSA : Triptone Soy Agar
UV : Ultraviyole
1. GĠRĠġ
Meme bezinin yangısı olarak tanımlanan mastitis, tüm dünyada süt yönlü yetiştiriciliğin en önemli problemidir. Modern işletmelerde uygulanan kontrol programlarına rağmen önemli ekonomik kayıplara neden olmaktadır (Aydın ve Paracıkoğlu, 2006; Quinn ve ark., 1994; Tel ve ark., 2009).
Tüm evcil hayvanlarda görülmesine rağmen özellikle süt ineklerinde önemli bir sorun olduğu bilinmektedir. Ülkemizde mastitis üzerine birçok çalışma yapılmasına rağmen halen süt sığırı yetiştiriciliğinde yaygın olarak görülmektedir (Tel ve ark., 2009). Subklinik mastitisler, klinik mastitis olgularına göre daha yaygın olarak bulunmakta ve daha fazla ekonomik kayba neden olmaktadır (Philpot ve Nickerson, 1991; Tel ve ark., 2009). Meme bezinin yangısı olarak tanımlanan mastitis, süt inekçiliğinde en yaygın olarak görülen ve en fazla ekonomik kayba neden olan bir hastalıktır (Gürbulak ve ark., 2009).
Mastitis, etkenlerin bulaşma şekilleri, korunma ve tedavisi bakımından oldukça karmaşık bir yapı göstermektedir (Harmon, 1994; Kaya ve Güven, 2008). Hastalık süt veriminin düşmesine, sütün bileşiminin değişmesine ve ineklerin üretken ömürlerinin kısalmasına neden olmaktadır. Mastitise bağlı süt kayıplarının yaklaşık %70-80’i subklinik mastitisten kaynaklanmaktadır (Jansen, 1970).
Mastitisler, memenin parankim dokusu, süt kanalları ve interstisiyel dokusunun yangısı olarak bilinir. Mastitisler dünyanın birçok ülkesinde ve ülkemizde de görülmektedir (Alaçam ve ark., 1989; Pir ve ark., 1983; Gülcü ve Ertaş, 2004). Mastitis olgularında birçok etken izole ve identifiye edilmiştir (Pitkala ve ark., 2004; Watts ve ark., 1995). Bu etkenlerin çok önemli bir kısmını bakteriler oluşturmaktadır (Tel ve ark., 2009).
Mastitisler meme dokusunda gelişen yangının şiddetine göre klinik ve subklinik tarzda görülür. Klinik mastitisler, meme dokusunda ve sütte gözle görülebilir değişikliklerle karakterizedir. Ayrıca klinik mastitisler perakut, akut, subakut ve kronik mastitis olarak da sınıflandırılmaktadır. Perakut formun prognozu çoğunlukla olumsuz olmasına rağmen, kronik subklinik form en önemli ekonomik kayıpların görüldüğü formdur (Baştan, 2002). Perakut form, genellikle doğumu takiben kısa sürede ortaya çıkar ve mortalitesi yüksektir. Süt sulu, kanlı, flakonlu ve pıhtılı görünümde olabilir (Baştan, 2002). Yüksek ateş, hızlı kalp atışı, anoreksi, depresyon, rumen hareketlerinde azalma ve kaslarda güçsüzlük gibi şiddetli sistemik reaksiyonlar gözlenebilir. Lokal ve
sistemik reaksiyonlar ani olarak gelişir. Normal gözüken inek aniden yatabilir ve komaya girebilir. Memeler aşırı şişmiş, sert ve ağrılıdır. Meme ucunda başlayan morarma daha sonra meme loplarını da kapsayabilir. Etkilenmeyen meme lopları da genellikle şişmiştir. Tokseminin hızlı geliştiği vakalar ölümle sonuçlanır (Akay ve ark., 1984; Baştan, 2002). Akut mastitisler, genellikle laktasyonun erken dönemlerinde meydana gelir. Memelerde şiddetli şişkinlik, sütte irin veya kan pıhtıları görülür. Akut formda sistemik belirtiler olarak ateş, anoreksi, depresyon, zayıflama görülür. Lokal semptomlar ise meme bölgesinde şişme, ağrı, ödem ve sıcaklık artışı gibi yangısal belirtiler ile karakterizedir (Karahan, 2005). En yaygın görülen mastitis formu subklinik formdur. Subklinik formun başlangıcında memede ve hayvanın genel durumunda bir değişiklik farkedilmez. Fakat ilerleyen dönemlerde sütün yapısında değişiklikler meydana gelir. Sütte sulu bir görünüm veya pıhtı oluşumu ile birlikte, memelerde atrofi ve yavaş şekilde gelişen sertleşmeler gözlenebilir. Sütte somatik hücre sayısı artmıştır. Sütte meydana gelen değişiklikleri indirekt saptayan testler veya memelerin palpasyonu düzenli şekilde yapılmadığı takdirde hastalık memelerin fonksiyonlarını kaybetmesi ile sonuçlanabilir (Akay ve Aydın, 1984).
Mastitisin sebepleri hazırlayıcı ve yapıcı sebepler olmak üzere ikiye ayrılır. Yapıcı sebeplerin en önemlileri mikroorganizmalardır. Mastitis olgularında çeşitli mikroorganizma cinslerine ait birçok etken izole ve identifiye edilmiştir. İnek mastitislerinde Staphylococcus aureus (S. aureus), Staphylococcus epidermidis,
Streptococcus agalactiae, Streptococcus dysgalactiae, Streptococcus uberis, Actinomyces pyogenes ve koliform gurubu bakteriler en sık izole ve identifiye edilen
etkenler olarak bildirilmiştir (Alaçam ve ark., 1989; Aydın ve Coşkuner, 1983). Daha seyrek olarak da Corynebacterium bovis, Bacillus subtilis, Bacillus cereus, Micrococcus
spp., Candida albicans, Cryptococcus spp., Nocardia spp. gibi çeşlitli bakteri ve
mantarların etkili oldukları bildirilmektedir (Alaçam ve ark., 1989; Ateş ve ark., 1991; Aydın ve Akay, 1984; Erer ve ark., 1996; Gülcü ve Ertaş, 2004).
Gülcü ve Ertaş (2004) yaptıkları çalışmada 160 ineğe ait mastitisli 206 meme lobundan etken izolasyonu yapmışlardır. Bunlardan Staphylococcus aureus %39.04,
Staphylococcus epidermidis %17.81, Actinomyces pyogenes %14.38, Escherichia coli
%8.9, Streptococcus agalactiae %6.16, Streptococcus uberis %4.11, Streptococcus
dysgalactiae %3.42, Corynebacterium bovis %1.37, Pasteurella haemolytica %1.37 ve Bacillus cereus %1.37 oranında izole etmişlerdir.
Özdemir (2005) mastitisli inek sütlerinden izole edilen stafilokok cinsine ait türleri identifiye ederek, bölgedeki dominant stafilokok türleri hakkında epidemiyolojik veriler elde etmek amacıyla bir çalışma yapmıştır. Bu çalışmada, mastitis bulguları gösteren vakalardan alınan toplam 448 süt örneğinden Micrococcaceae familyasına ait katalaz ve Gram pozitif kok olduğu saptanan 292 bakteri, kültürel ve biyokimyasal özelliklerine göre identifiye edilmiştir. Yapılan testler sonucunda, incelenen 292 izolatın 25 (%8.56)’i Micrococcus spp, 154 (%52.7)’ü Staphylococcus aureus, 48 (%16.4)’i S.
epidermidis, 32 (%10.9)’si S. cohnii, 21 (%7.2)’i S. saprophyticus ve 12 (%4.1)’si S. hyicus olarak identifiye edilmiştir. Stafilokok cinsine ait 267 izolatın 164’ü koagülaz
pozitif (%61.5) ve 103’ü koagülaz negatif (%38.5) olarak bulunmuş. Koagülaz pozitif suşların 154 (%94)’ü S. aureus, 10 (%6)’u S. hyicus olarak belirlenirken, koagülaz negatif suşların 48 (%46.6)’i S. epidermidis, 32 (%31.1)’si S. cohnii, 21 (%20.3)’i S.
saprophyticus ve 2 (%1.9)’si de S. hyicus olarak identifiye edilmiştir.
Staphylococcus cinsi içerisinde en az 35 tür yer almaktadır. Bu cins içinde en çok bilineni S. aureus’tur. Diğer türlerden farklı olarak S. aureus, koagülaz pozitiftir ve en önemli patojendir. Hemen herkes hayatları süresince S. aureus enfeksiyonlarının bazı tipleri ile karşılaşmaktadır. Geçmişte sadece normal flora bakterisi olarak kabul edilen koagülaz negatif stafilokok (KNS) türlerinin günümüzde zararsız etkenler olmadığı bilinmektedir (Taponen ve Pyorola, 2009).
S. aureus’un en önemli yüzey proteinlerinden biri olan spa hücre duvarının
%7’sini teşkil eder. Bu protein IgG’nin Fc bölgesine bağlanarak opsonofagositozu inhibe eder. Bu bağlanma aynı zamanda hem klasik hem de alternatif yolla komplementin aktivasyonuna neden olur (Palmqvist ve ark., 2002; Gao ark., 2004; Todar, 2005; Erdem, 2011). Spa; S. aureus suşlarınınn %90-95’inde bulunur ve sadece
S. aureus suşlarında bulunan gruba özgü bir antijendir (Akan, 1993; Todar, 2005;
Erdem, 2011).
Koagülaz, fibrinojeni fibrine çevirme özelliğinde protrombin benzeri aktiviteye sahip bir proteindir. Koagülaz enzimi enfeksiyon sırasında in vivo fibrin bir bariyer oluşturur ve bakteriyi fagositozdan korur. Koagülaz bağlı ve serbest olmak üzere iki formda bulunur. Bağlı koagülaz (clumping factor) bakteriyel hücre duvarına bağlı halde bulunur, kültür filtratlarında serbest olarak bulunmaz. Bakteriler plazma içinde süspanse edildiğinde, bakteriler arasında fibrin iplikçikleri oluşur ve bakterilerin gözle görünen kümeler oluşturmasına neden olur. Serbest koagülaz ise kültür filtratlarında bulunan bir
enzimdir. Koagülaz üreten mikroorganizma süspansiyonu plazma ile bir tüpte karıştırıldığında görülebilir pıhtı oluşur (Koneman, 1997).
Klamping faktör A (clfA), S. aureus’un konakçı hücreye tutunması için en önemli adezyon faktörlerinden biridir. ClfA, çözünebilir veya immobilize fibrinojen ile bakteriyel bağlanmayı kolaylaştırarak enfeksiyon sürecine katılmaktadır. Hayvan enfeksiyonlarında clfA’nın polimorf nüklear lökositlerin bakteriyi fagositozunu engelleme yeteneği önemlidir (Mcdevit ve ark., 1994; Karahan ve ark., 2005). S. aureus patogenezinde rol oynayan moleküler mekanizmaların detaylı incelenmesi ve belirlenmesi gelecekteki daha etkili profilaktik tedavi uygulamalarında dikkate alınması önemlidir (Klein ve ark., 2012). Fenotipik ve genotipik karakterizasyon sığır mastitis izolatları arasında S. aureus klonlarının sıklığının ve dağılımının daha iyi anlaşılmasını sağlayabilir. Bu incelemeler süt ve sürülerde S. aureus enfeksiyonlarının kontrolünde yardımcı olabilir (Kalorey ve ark., 2007). S. aureus sığır mastitisinin önde gelen güçlü bir patojenidir. Enfeksiyonun başarısız olması için bir dizi bakteriyel proteinleri tanımlamaya çalışmak çok önemlidir. Bu proteinlerin tanımlanması potansiyel aşı hedeflerinin tespiti amaçlı kullanılması için önemlidir (Klein ve ark., 2012).
Bu çalışmada mastitisli ineklerin sütlerinden izole edilmiş olan S. aureus suşlarında bakterinin patogenezisine katkıda bulunan bazı virulans faktörlerin belirlenmesi ve tiplendirilmesi amaçlanmıştır.
2. KAYNAK ARAġTIRMASI
Süt sığırlarında yaygın olarak görülen hastalıklardan biri olan mastitis meme bezleri dokusunun yangısal bir yanıtıdır. Mastitis süt endüstrisi için önemli bir kayba neden olur. Mastitise neden olabilen çeşitli bakteriyel çeşitli bakteriyel patojenler arasında S. aureus, meme bezlerinde tedavi edilmesi son derece güç olan kronik ve derin enfeksiyonlara neden muhtemelen en öldürücü etkenidir (Momtaz ve ark., 2010).
Mastitis, tüm dünyada süt yönlü yetiştiriciliğin en önemli problemidir. Modern işletmelerde uygulanan kontrol programlarına rağmen önemli ekonomik kayıplara neden olmaktadır (Tel ve ark., 2009). Ülkemizde ineklerde mastitis görülme oranının %30 dolayında olduğu ve mastitis nedeniyle süt veriminde yaklaşık %10 oranında azalma meydana geldiği, bunun sonucunda da yıllık ekonomik kaybın trilyonlarla ifade edilebileceği bildirilmektedir (Mutluer, 2001; Sabuncuoğlu ve ark., 2003).
Enfeksiyon; bakteri, maya, mantar ve virüs gibi bulaşıcı etmenler ile stres, yaralanmalar, yüksek kalorili yemler, ineğe ve çevreye bağlı pek çok faktörler nedeniyle meydana gelmektedir. Enfeksiyonun sonucunda sütteki kan proteinlerinin ve beyaz kan hücrelerinin sayısı artmaktadır (Erdem ve Atasever, 2004). Enfeksiyona yol açan mikroorganizmaların yaklaşık %95’ini Streptococcus agalactiae, Staphylococcus
aureus, Streptococcus dysgalactiae, Streptococcus uberis ve Escherichia coli, %5’ini
ise diğer mikroorganizmalar oluşturmaktadır (Bray ve Shearer, 1994). İneklerde mastitisin ortaya çıkmasındaki hazırlayıcı faktörler; ineğe ilişkin olanlar, bakım ve çevre faktörleri ile sağım süreci olarak gruplandırılabilir. İneğe bağlı olanlar; ırk ve kalıtım, yaş ve laktasyon sayısı, meme yapısı, süt verimi, laktasyon dönemi, immun sistemin gücü, meme ve meme başındaki lezyonlar ile sağım kolaylığı olarak sıralanabilir. Bakım ve çevre unsurları; mikroorganizmalar, çevre, ahır düzeni ve koşulları, zemin ve altlıklar, bağlanma şekli, beslenme, egzersiz ve mevsimin etkisi olarak değerlendirilebilir. Sağım süreci ise; sağım öncesi uyarma, sağım hijyeni ve tekniği, sağım makinesinin sorunları ve sağımcının deneyimi gibi faktörlerden oluşmaktadır (Balık, 1998).
Sığır varlığı bakımından AB ülkeleri içinde üçüncü, dünyada ise 27. sırada olan ülkemizde süt üretimi içinde sığırın payı her geçen yıl artış göstermektedir. 2005 yılı verilerine göre yaklaşık 4 milyon baş sağmal inekten 10 milyon ton süt elde edilmekte, bu miktar toplam üretilen sütün %90.26’sını oluşturmaktadır. Buna karşın, bugün Türkiye’de, çoğu ülkedeki gibi tarımsal ekonominin geniş ve dinamik bir iş kolunu
oluşturan süt endüstrisinin ve süt sığırı yetiştiricilerinin karşı karşıya kaldığı sorunların en başında Mastitis gelmektedir (Atasever ve Erdem, 2008).
Mastitisler, meme dokusunda ve sütte oluşan patolojik, fiziksel, kimyasal ve bakteriyolojik değişikliklere göre klinik ve subklinik mastitis olarak sınıflandırılır. Klinik mastitis, sütte ve meme dokusunda önemli yangısal belirtiler ile karakterize olan mastitis formudur. Subklinik mastitislerde ise memede ve sütte gözle görülür değişiklikler oluşmamaktadır. Bu nedenle subklinik mastitisler uzun süre devam etmekte ve daha fazla ekonomik kayıplara neden olmaktadır. Subklinik mastitisler, klinik mastitislere oranla daha yaygın görülmektedir. Ortalama olarak her klinik mastitise karşılık 20-40 subklinik mastitis olgusuyla karşılaşılmaktadır. Bu olgularda ekonomik kayıp, zarar gören dokular sebebiyle süt üretiminde %8’e varan azalma nedeniyledir. Mastitis olgularından birçok etken izole ve identifiye edilmiştir (Macun, 2011).
Mastitisli sütlerden izole edilen stafilokokların koagülaz pozitifliğine bakılarak bakterinin S. aureus olarak isimlendirilmesi birçok mikrobiyoloji laboratuvarı tarafından yeterli olarak görülmektedir. Ancak S. aureus dışında koagülaz pozitif özellik gösteren stafilokok türlerinin de varlığı göz önüne alındığında koagülaz testinin dışında S. intermedius ve S. hyicus gibi koagülaz pozitif türlerin S. aureus’tan ayrımı için DNase ve klamping faktör testlerinin de yapılmasının gerekliliği vurgulanmıştır (Yavuz ve ark 2002). Stafilokokların laboratuvarlarda hızlı identifikasyonuna yönelik ticari StaphTrac, Minitek Gram-Positive Set ve Api-STAPH gibi kitler geliştirilmiştir (Goh ve ark., 1992; Tanrıbuyurdu, 2014).
Konakçı ile mikroorganizmaların bu ilişkisinde virulansta önemli bir paya sahip olan ve ekzopolisakkarid içeren dış tabakanın kuvvetlice bağlanma özelliğinin önemli bir rolü vardır. Slime, bir bakterinin uygun bir ortamda üremesinden sonra şekillenen ekzopolisakkarid bir matrikstir. Bir stafilokok suşunun slime üretebilen varyantlarının, üretmeyen varyantlarından daha yüksek derecede kolonizasyon yeteneğine sahip olduğu bildirilmiştir (Baselga ve ark., 1993; Arslan ve Uçan, 2005).
S. aureus insanlarda ve hayvanlarda çok ciddi hastalıklara yol açan bir patojen
olarak dünya çapında tanınan ve klinik ve subklinik mastitisin sığırlarda en sık rastlanan etkenlerinden biridir (Yang ve ark., 2012). S. aureus suşları doğal olarak ineklerin derisinde ve muköz epitelyum hücrelerinde yaşamalarına rağmen inek meme bezlerini infekte ederek mastitise de yol açabilmektedir. Bu süreç hücresel bağlanma ve invazyonu kolaylaştırması yanı sıra konakçı immun yanıtları ve toksik doku
reaksiyonlarından kurtulmasını kolaylaştıran hüre yüzeyindeki çeşitli virulans faktörlerini içermektedir (Kerro Dego ve ark., 2002; Stutz ve ark., 2011). S. aureus mastitis de dâhil olmak üzere ineklerde çok çeşitli hastalıklara yol açan, virulans faktör olarak görev yapan birçok proteini kodlar. Bu virulans faktörlerin arasında, spa, clfA, clfB, FnbA, ve FnbB konak hücrelerin işgalinin yanı sıra, konak bağışıklık yanıtlarından kaçınmak için de önemlidir (Stutz ve ark., 2011). S. aureus konakta enfeksiyon oluştururken yapısında bulunan birçok yapısal elemanlardan, enzimlerinden ve toksinlerinden yararlanır. Bu patojenite faktörleri sayesinde fagositozdan kaçabilir, deri ve deri altına kolayca yayılabilir ve tipik apse formunu oluşturabilirler. Ayrıca toksinleri aracılığı ile farklı enfeksiyonlara da neden olurlar (Koneman, 1997).
Klinik örneklerden en sık izole edilen stafilokok türü deride flora etkeni olan
Staphylococcus epidermidis’tir (Kawamura ve ark., 1998; Alen ve ark., 2006).
Stafilokoklar genelde bulundukları yerde konakla iyi huylu ve simbiyotik bir ilişkiye sahiptirler, ancak deri ve mukozada travma, enjeksiyon veya cerrahi müdahaleler sonucu oluşan portantreler ile dokuya girerek patojen olabilirler (Bannerman, 2003; Erdem, 2011). S. aureus insan ve hayvanlarda çok sayıda hastalığa neden olabilmektedirler. İneklerde görülen mastitislerin en önemli patojenlerinden biridir (Akan, 2006; Ünal, 2013). Mastitise neden olan birçok mikroorganizma ise insanlar için de sakıncalıdır. Pastörizasyon ile insanda hastalık yapan bu mikroorganizmaların çoğu yok edilebilmektedir. Ancak bazı stafilokok suşları, sütte toksin üretirler ve bu toksinler kaynatma ve kurutmaya dayanıklıdır, sindirim sisteminde irritasyonla ishal ve kusmaya neden olurlar. Dolayısıyla S. aureus’un neden olduğu mastitislerin tedavi edilmesi hem hayvan sağlığı hem de insan sağlığı açısından büyük önem taşımaktadır (Deveci ve ark., 1994; Alaçam ve ark., 1986).
S. aureus’un başlangıçtaki bağlanması miyoepitel hücreler ve fibroblastlar
etrafındaki lokalize olan, fibronektin bağlayıcı proteinler A ve B (FnbA ve FnbB) ve klamping faktör (clfA ve clfB) gibi bakteri yüzey proteinlerinin etkileşimine bağlıdır. Ayrıca fibronektin-bağlayıcı proteinlerin, sığır meme bezi hücrelerine yapışma ve işgal etme rollerinin olduğu gösterilmiştir (Asch ve ark., 1981; John ve ark., 1999; Sutra ve Poutrel, 1994; Stutz ve ark., 2010). Fibrinojen bağlayan MSCRAMM’lar (ClfA- ClfB)
S. aureus’un fibrinojen kaplı substratlara bağlanma ve fibrinojen varlığında aglütine
olabilme yeteneğini sağlayan bu protein, kümelenme faktörü (Clumping Factor–ClfA) adını alır (Foster ve Höök, 1998). Bağlı koagülaz olarak da adlandırılan kümelenme faktörü Stafilokokların hücre yüzeyinde meydana gelir ve serbest bırakılmaz (Akan,
1993). S. aureus’ un fibrinojene spesifik olarak bağlanması temel olarak clfA ve clfB ile sağlanır. ClfA’nın inaktivasyonu, in vitro ve in vivo koşullarda fibrinojen kaplı yüzeylere S. aureus’ un bağlanmasını inhibe eder (Wolz ve ark., 2002). ClfA bakterinin tüm çoğalma dönemlerinde bulunmasına rağmen, clfB sadece erken çoğalma fazındaki hücrelerde bulunur ve durağan fazdaki hücrelerin yüzeyinden kaybolur (Foster ve Höök, 1998; Erdem, 2011).
S. aureus’da fibronektin bağlayıcı proteinler sığır meme bezinin bakteriyel
yapışmasında ve işgaline katkıda bulunan önemli virulans faktörleridir. S. aureus’da iki fibronektin bağlayıcı protein olarak fnbA ve fnbB tanımlanmıştır ve onlara karşılık gelen genlerin dizi benzerliği yüksektir (Proietti ve ark., 2010).
S. aureus hücre duvarında bulunan gruba özel bir antijen olan protein A Wervey
tarafından 1940’da tanımlanmıştır. Molekül ağırlığı 13000 dalton olan küçük bir proteindir stafilokoklarda üç tip spa bulunmuştur. Bunlar serbest protein A, hücreye bağlı protein A ve hücre dışı protein A olarak isimlendirilir. Üreme sırasında besiyerine salgılanan protein A bakterinin fagasitozunu önlemektedir. Protein A, S. aureus’un hücre duvarı kompenentlerinden birisi olup, büyük bir kısmı peptidoglikan yapıya kovalan olarak bağlanmıştır. Bir kısmı ise hücre dışı ortama salınmaktadır. Spa komplemanı aktive eder, antifagositik, kemotaktik, mitojenik etkileri vardır. Spa’nın koagülaz ve nükleaz aktiviteleri ile büyük oranda korelasyon göstermesi, antifagositik etki yaratması ve antibiyotik duyarlılığını azaltması gibi özellikleri patojenite kriteri olacağına işaret etmektedir (Demirpek, 2006). Çoğu S. aureus suşlarının yüzeyi peptidoglikan ya da sitoplazmik membrana bağlanan protein A ile kaplanmıştır (Iraz, 2008). Bu yüzey proteinleri ektraselüler doku proteinlerine bağlanarak solid yüzeylere aderansın başlamasına da anahtar rol oynamaktadır (Şahin, 2007). Bu protein bir polimorfik (X) ve bir de korunmuş bir bölge içeren spa geni tarafından kodlanmaktadır. Polimorfik X bölgesi çeşitli sayıda 24 bç’lık tekrarlar içermektedir (Koreen ve ark., 2004; Coelho ve ark., 2009). Protein A, S. aureus hücre duvarında IgG’nin Fc bölgesine bağlanma özelliğinde olan bir proteindir. Hücre duvarına bağlı olarak bulunabildiği gibi üreme sırasında besi yeri ortamına da salgılanabilmektedir. Protein A’nın opsonizasyonu ve fagositozu engellemesi, kompleman aktive etmesi ve gecikmiş aşırı duyarlılık reaksiyonlarına neden olmasından dolayı bir virulans faktörü olarak fonksiyon görmektedir. Kapsül, bazı S. aureus suşlarının ürettiği bakteriyi fagositozdan koruyan ekzopolisakkarittir. Bu ekzopolisakkarit materyal aynı zamanda etkenin hücrelere ve prostetik cihazlara adherensini artırmaktadır. S. aureus’un klinik izolatları
kapsüler polisakkarit antijenlerine göre 8 serotipe klasifiye edilmiştir. En yaygın serotiplerin serotip 5 ve 8 olduğu bildirilmiştir (Koneman, 1997). Protein A’dan yoksun
S. aureus suşlarının, in vitro koşullarda kolayca fagosite olduğu gösterildiğinden spa
önemli bir virulans faktörü olarak kabul edilir (Akan, 1993; Todar, 2005; Erdem, 2011). Protein A IgG1, IgG2 ve IgG4'lerin Fc bölgelerine bağlanarak antikor bağımlı klirensi inhibe eder, lökosit kemotaksisini uyarır, anti-kompleman özelliktedir (Boz, 2009).
Koagülaz proteini, doğal pıhtılaşmadan farklı olan bir mekanizma ile fibrinojeni
fibrin ipliklerine dönüştürme yeteneğine sahiptir (Palma ve ark., 1999; Coelho ve ark., 2009). Koagülaz meme bezi enfeksiyonunda bir virulans faktör olarak rol oynamaktadır. Bu protein S. aureus izolatları arasında akrabalık ilişkisini belirlemek için kullanılabilen bir tekrarlı polimorfik bölgeye ve bir de korunmuş bölgeye sahip olan coa geni tarafından kodlanır. Bu bölge dizisi ve sayısı değişken 81 değişken baz çiftlik tekrar dizileri ve 330 baz çiftlik korunmuş dizileri içermektedir (Shopsin ve ark., 2000; Reinoso ve ark., 2004; Coelho ve ark., 2009). Koagülaz Serbest ve bağlı olmak üzere iki tipi vardır; Serbest koagülaz: Ekstrasellüler salgılanabilen bir proenzim olan koagülaz plazmadaki CRF (coagulase-reacting factor) ile birleşerek aktif duruma geçer ve fibrinojenin fibrine dönüşümünü uyararak plazmayı pıhtılaştırır. Antijenik olarak 4 farklı tipi vardır. Filtrelerden geçebilir ve ısıya dirençli bir enzimdir. Bağlı koagülaz (Clumping Factor): Hücre duvarına bağlı halde fibrinojeni fibrine çevirerek hücre yüzeyinde fibrin presipitasyonu meydana getirir. Bunun sonucu olarak stafilokoklar aglütinasyon ve kümeleşmeye uğrar. Bakterinin patojenitesindeki rolü, fagositozu önlemek üzere bakterinin üzerine fibrin tabakası örtmesinden ileri gelmektedir. Bunlardan başka stafilokokal serin proteaz (SspA), sistein proteaz (SspP), metalloproteaz (aureolysin; Aur) ve staphopain (Scp) gibi ekstra sellüler proteazlar da sentezlerler (Boz, 2009).
Karahan ve ark. (2011) Türkiye’deki mastitisli ineklerden izole edilen, coa pozitif S. aureus suşlarında bulunan, klamping faktör (clfA) ve protein A (spa) genlerini PCR tekniği kullanarak, karakterizasyonunu yapmışlardır. Bu çalışmada, coa pozitif suşların hepsinde, X ve IgG bağlayıcı bölgeleri kodlayan spa geninin varlığını tespit etmişlerdir. Diğer taraftan, 84 (%91.3) izolatda clfA genini içerdiğini gözlemlemişlerdir. Sığır kökenli S. aureus izolatlarının tiplendirilmesinde spa gen polimorfizmleri ve coa PCR birlikte kullanıldığında geleneksel yöntemlere göre iyi bir alternatif olarak önerilebileceği sonucuna varmışlardır. Bu da stafilokokal mastitise karşı etkili kontrol yöntemlerinin geliştirilmesi için önemli bilgiler sağlamaktadır. Sonuç olarak,
Türkiye’de mastitise yol açan S. aureus suşlarının genetik olarak çeşitlilik gösterdiği kanıtlanmıştır. Türkiye‘nin doğusundaki mastitisli ineklerden izole edilmiş olan coa pozitif S. aureus suşlarında önemli virulans faktör olan clfA, spa-IgG ve spa-X, genlerinin mevcut olduğunu gözlemlemişlerdir.
Coelho ve ark. (2008) çalışmalarında, subklinik mastitisli ineklere ait sütlerden izole edilmiş S. aureus’da direnç paterni ile virulans faktörlerin fenotipik ve genotipik olarak tanımlamayı amaçlamışlardır. Çalışmalarının sonuçlarına göre, coa geninin amplifikasyonu ile yaklaşık olarak 400 bç, 600 bç, 700 bç ve 900 bç uzunluğunda olmak üzere dört farklı polimorfizm tespit etmişlerdir. Protein-A IgG bağlanma bölgesi kodlayan spa geni 700 bç ve 900 bç'lik boyutlarda olduğunu göstermişlerdir. Ayrıca her bir izolatta, spa geninin X bölgesi için, yaygın olarak 180 bç’lik tek bir uzunluk elde etmişlerdir.
Akineden ve ark. (2001) yaptıkları çalışmada Almanya’nın bir bölgesinde 7 farklı yerdeki 8 farklı çiftlikten seçilen 60 mastitisli ineğin sütlerinden izole edilen 103 farklı S. aureus suşlarına ait çeşitli toksinlerin genotipik ve fenotipik tanımlarını yaparak karşılaştırmışlardır. S. aureus izolatları, bakterinin kültür ve hemolitik özellikleri ile tüpte koagülaz reaksiyonunun belirlenmesi esas alınarak, tanımlanabilir. 23S rRNA kodlayan genin türe özgü spesifik parçaları PCR amplifikasyonu ile doğrulanmıştır. Bununla birlikte izolatların hepsinin protein A da bulunan IgG bağlayıcı bölgeyi ve X bölgesini kodlayan genlerle, koagülaz ve klamping faktörü kodlayan genleri bulundurduğunu saptamışlardır.
Zecconi ve ark. (2006) yaptıkları çalışmada, koagülaz geni (coa), protein A geni (spa), kolajen bağlayıcı protein geni (CNA), fibrinojen bağlayıcı protein geni (EFP), Panton-Valentine lökosidin geni (pvl) ve enterotoksinler (sea, seb, sec, sed, see, seg,
seh, sei, sej) gibi çeşitli virulans faktörlerin kombinasyonunun meme yangısına cevapta,
farklı genlerin varlığı veya yokluğu arasındaki ilişkinin analiz edilmesini amaçlamışlardır. Virulans genlerin arasındaki bu ilişkinin analizi subklinik mastitisin oluşumunda spa geninin ve bir enteretoksin olan sej geninin risk faktörü olarak rol oynadığı gösterilmiştir. Bu sonuçlar aynı zamanda subklinik mastitisin gelişiminde, virulans gen kombinasyonları tarafından izolatların bir küme içinde sınıflandırılmasını sağlayan, suşların karakteristik özellikleri ve virulans faktörlerin spesifik kombinasyonlarının ekspresyonları ile ilişkili olabileceğini desteklemektedir.
Momtaz ve ark. (2010) mastitisli çiğ süt örneklerinden izole ettikleri 360 S.
aureus’da bazı virulans faktörleri özel primerler kullanarak PCR ile belirlemişlerdir. Bu
360 çiğ süt örnekleri arasında 86 suştan, 42 örneğin coa genini, 63 örneğin clfA genini, 69 örneğin protein A geninin X bölgesini kodlayan geni, 22 örneğin IgG bağlayıcı gen bölgesi içerdiğini rapor etmişlerdir. Bu çalışma ile clfA ve X bölgelerinin sentezinin patojenik S. aureus suşlarından en sık izole edilen genler olduğu gösterilmiştir.
El- Sayed ve ark. (2010) yaptıkları çalışmada mastitisli sığırlardan izole edilen S.
aureus suşlarında (M1, M2, M3, M4) protein A’yı kodlayan spa geni PCR ile
çoğaltılarak dizilenmiştir. 41 suştan 4’ü seçilerek detaylı incelenmiştir. M1, M2, M3 suşları IgA bağlayan domainlerini kodlayan gen segmentlerinden beş tanesini ( E, D, A, B ve C) içerirken, M2 suşunda bu gen segmentlerinden dört tanesini (E, A, B, C) içermektedir. Seçilmiş dört suşun (M1, M2, M3 ve M4), spa genin çoğaltılmasıyla elde edilen boyutlar sırasıyla 1050, 800, 1000 ve 1120 bç olduğu gözlenmiştir. IgG bağlayıcı bölgeyi kodlayan gen segmentleri ve X bölgesi için oligonükleotid primerler tasarlanmış spa 1, spa 2 ile Spa 3, Spa 4, IgG bağlayıcı bölgeyi kodlayan gen bölgelerinin amplikasyon boyutları sırasıyla; 900 bç (M1 ve M3), 800 bç (M4) ve 700 bç (M2) olarak gözlenmiştir. X bölgesini kodlayan segmentler sırasıyla; 320 bç (M4), 150 bç (M1) ve 100 bç (M2 ve M3) uzunluğunda olduğu belirtilmiştir. Bu çalışmada mastitisli sığırlardan izole edilen S. aureus’a ait spa geninde büyüklüklerinde ve dizilerinde yüksek çeşitlilik gözlenmiştir. Bütün dizi varyansların virulans üzerine etkisinin olduğu tespit edilmiştir.
Arjantin’de mastitisli ineklerden ve yiyeceklerden izole edilmiş 45 S. aureus suşunun genotipik olarak karakterize edilmesinin amaçlandığı çalışmada protein A, koagülaz, klamping faktör gibi bazı virulans faktörleri kodlayan genleri PCR tekniği ile analiz edilmiştir. IgG bağlanma bölgesinin 900 bç’lık ve 700 bç’lik büyüklüğünde iki farklı PCR amplifikasyonu elde edilmiştir. X bölgesinin büyüklüğünün 100 bç dan 315 bç’a kadar oldukça polimorfizm gösterdiğini, coa geninin amplifikasyonu sonucunda 7 farklı tip gözlendiğini ve clfA geninin ise 1000 bç’lik bir amplikon gözlediklerini belirtmişlerdir (Reinoso ve ark., 2008).
Stephan ve ark. (2001) enterotoksin üreten mastitisli ineklerden izole edilen suşlardan 34 S. aureus suşunda bazı virulans genlerinin belirlenmesini amaçlamışlardır. Çalışmanın sonunda bütün suşlarda clfA’nın 980 bç büyüklüğünde tek bir amplikon gözlediklerini, spa geninin X bölgesinin ise 4 farklı büyüklüklerde her bir suşta tek bir amplikon gözlediklerini ve farklı büyüklüklerin 2, 6, 10 ve 11 tekrarlardan oluştuğunu
bildirmişlerdir. coa geni amplifikasyon sonunda da 580-660 bç büyüklüklerinde her bir suşta bir amplikon gözlemişlerdir. Bu çalışma yakın akraba S. aureus klonlarının mastitisten sorumlu olduğunu göstermiştir.
Khan ve ark. (2013) mastitisli sığır ve mandalardan izole ettikleri S. aureus’ların genotipik özelliklerini ve dağılımını belirlemek amacıyla çalışma yapmışlardır. Süt veriminin düşük olması nedeniyle Faisalabad ilçe yakınlarındaki iki deneysel hayvan çiftlikleri olan Okara (Bahadar Nagar) ve Sahiwal (Qadirabad)’da, hazır bulunan ve bir mezbahada kesilen, emzikli sığır (653) ve manda (792) olmak üzere toplam 1445 hayvanı mastitis bakımından taramışlardır. Mastitisi tespit etmek amacıyla CMT (Kaliforniya Mastitis Testi) kullanmışlardır. Pozitif süt örneklerinin çeyreğini S. aureus izolatlarının kültürü için toplamışlar. S. aureus izolatlarının, büyüme özellikleri, biyokimyasal özellikleri, koagülaz testi ve yanı sıra koagülaz (coa) ve spa (spa-X) gibi virulans genlerini çalışmışlardır. S. aureus izolatları (n=265) koagülaz (coa) ve spa (spa-X) genlerin sıklığını belirlemek için polimeraz zincir reaksiyonu (PCR) ile karakterize etmişlerdir. Bu izolatlardan spa-X, geni boyutunda 190 bç ile 320 bç arasında değişen beş farklı büyüklük, coa geninde ise 490 bç ile 204 bç büyüklüklerinde PCR ürünleri elde etmişlerdir. Buna göre tüm S. aureus suşlarının koagülaz pozitif olduğu, 261 (%98.5) izolatta ise spa-X geninin varlığı bulunmuştur.
Algammal ve ark. (2013) Subklinal mastitisli ineklerin sütlerinden geleneksel yöntemlerle izole edilmiş 84 S. aureus izolatlarında virulans gen faktörlerini ve farklı ekzotoksinleri belirlemek amacıyla yaptıkları çalışmada 28 S. aureus izolatında koagülaz geni, protein A (spa) geninin X bölgesi ve pvl geninin tespiti için PCR yapmışlardır. Bu izolatların tamamında (%100) koagülaz geni, 26 (%92.8) izolatta farklı büyüklükteki spa-X geni, sadece 3 (%10.7) izolatta pvl geni bulunduğunu gözlemlemişlerdir.
Kalorey ve ark. (2007) çalışmalarında mastitisli sığırlardan izole edilen S.
aureus‘ların genotipik özelliklerini belirlemek amacıyla S. aureus suşlarının fenotipik
karakterizazyonunun yanında, koagülaz, clamping faktörü, thermonuclease (nuc), enterotoxin A (entA), spa geninin immunoglobulin G bağlayan bölgesi ve X bölgesi pcr ile genetik karakterizasyonunu yapmışlardır. İzolatların hepsi clfA genleri yaklaşık olarak 1,042 bç kadar bir boyutu olan amplikon vermiştir. Spa-IgG geni 34, spa-X geni de 26 izolatta saptamışlardır. Coa geninin 20, 10 ve 7 örnekte üç farklı boyutta, nuc genini 37 izolattan 36’sında gözlemlemişlerdir. Süt örneklerinde CMT kullanılarak mastitis belirlemişlerdir. 8 çiftlikteki 140 inekten toplanan 552 süt örneğinden, 134
inekteki 501 (%90.76) örnek pozitif CMT olarak bulundu. 114 inek ve bunlardan alınan 268 süt örneğinde, Staphylococcus sp. bulunduğunu gözlemlenmiştir. Kültürel ve biyokimyasal özellikleri dayanarak 37 izolat S. aureus olarak tespit edilmiştir. Tüm 37 izolatın koagülaz testi pozitif olarak bulunmuş. Diğer suşlar S. intermedius, S. hyicus, koagülaz negatif staphylacocci ve Micrococcus olarak tespit edilmiştir. Buna göre coa geninin amplifikasyonun da üç farklı büyüklükte ürün gözlenmiştir bunlar; 20 izolatta 627 bç, 10 izolatta 710 bç, 7 izolatta 910 bç şekindedir. Bütün izolatlarda clfA geninin yaklaşık olarak 1.042 bç., protein A’nın IgG bağlanma bölgesini kodlayan gen bölümünde; 12 izolatta 590 bç, 15 izolatta 810 bç ve 7 izolatta 970 bç, Spa geninin X bağlanma bölümü, 10 izolatta 220 bç, 9 izolatta 253 amplikon ve 7 izolatta 315 bç, nuc geninde 37 izolattan 36’sında 279 bç büyüklüğünde amplikon gözlemişlerdir. Örneklerin hiçbirinde entA geni bulunmadığı görülmüştür.
Kumar ve ark. (2011) mastitisli sığırlardan izole edilmiş S. aureus suşlarında çeşitli virulans faktörlerin ve toksinlerin genotipik ve moleküler varyasyonlarını çalışmışlardır. Biyokimyasal özelliklerine dayanarak, izolatların 128’i S. aureus olarak belirlemişlerdir. Bu izolatların, 83’ünü subklinik ve 45’ini klinik mastitis olarak gözlemlemişlerdir. subklinik ve klinik mastitisli suşlarda sırasıyla clfA geni %93.3, %87.9 ve coa geni %93.3, %91.6 oranlarında görülürken spa-X ve IgG bölgeleri 128 (%100) suşta da görülmüştür. Spa-IgG geninde; 500 bç, 1000 bç, 1050 bç, clfA geninde; 900 bç ve 1000 bç büyüklüklerinde bantlar bulmuşlardır.
Yang ve ark. (2012) yaptıkları çalışmada, Subklinik mastitisli ineklerden elde edilen S. aureus (n=39) suşlarında clfA, nbA, fnbB, cap5, cap8, hla, hlb, nuc, sea, ve tst kodlayan genlerin varlığını pcr ile belirlemeyi amaçlamışlardır. PCR sonuçlarına göre;
clfA 24 (%62), fnbA 38 (%97), fnbB 0 (%0), cap5 23 (%59), cap8 9 (%23), hla 33
(%85), hlb 32 (%82), nuc 35 (%90), sea 0 (%0), tst 1 (%3) izolatta olmak üzere bu genlerin varlığını belirlemişlerdir.
Başka bir çalışmada Brezilya Bahia’da süt örneklerinden izole edilen S. aureus suşlarının iltihap oluşturma kapasitesi üzerinde biyofilm oluşumu, virulans genlerin varlığı ve dağılımını incelemişlerdir. Vitória da Conquista belediyesinde, güneybatı Bahia da bulunan dört süt çiftliklerinden toplanan 132 Çiğ süt örnekleri üzerinde çalışma yapmışlardır. Örneklerin, 94’ünde (%71.2) S. aureus izolatı elde edilmiştir. Bu suşlar antibiyogramın yöntemine MIC (Minimum Inhibitory Concentration) tabi tutulmuş. Hastalık oluşturma özellikleri açısından glikozun neden olduğu in vitro biyofilm oluşum testleri yapmışlardır. izolatlar, Sea (enterotoksin A), seb (B), sec (C),
pvl, clfA ve spa tespiti için PCR ile genotiplendirilmiştir. 1 (%1.06) seb geni, 1 (%1.06) sec geni, 18 (%19.2) clfA geni ve 44 (%46.8) spa geni varlığını tespit etmişlerdir.
İzolatların hiçbirinde sea ve pvl geni olmadığı gözlenmiştir (Santos ve ark., 2014) Klein ve ark. (2011) yaptıkları çalışmada, mastitisli büyükbaş hayvanlardan elde edilmiş 85 bakteriyel izolatlattan yedi virulans faktörü kodlayan genin analizi yapılmıştır. En sık görülen ClfB geni %91.8, spa geni %85.9, sdrCDE geni %85.9,
fnBÇ geni %63.5, SSPA geni %62.4, clfA geni %50.6 ve cna geni %22.4 oranlarında
bulunurken en az bir genin bütün suşlarda bulunduğu ve bu genlerden birlikte bulunma bakımından en sık bulunan kombinasyonlar ise clfB ve sdrCDE (%82.4), clfB ve spa (%80.0) olduğu gözlenmiştir.
Proietti ve ark (2010), yaptıkları çalışmayla, İtalya'da yedi süt çiftliğinden toplanan sığır sütlerinden izole edilen S. aureus izolatının (n=170) genotipik özelliklerini incelemişlerdir. Genotipik karakterizasyon için, S. aureus izolatlarının virulans özellikleri kodlayan genleri (nuc, clfA, spa-IgG, spa-X, fnbA and fnbB, cap5 and cap8) ve stafilokokal enterotoksinleri (sea, seb, sec, sed, see, seg, seh, sei, sej) PCR tekniği kullanılarak incelemişlerdir. ClfA gen izolatlarının %82.4 oranında ve 1042 bç büyüklüğünde tespit etmişlerdir. Spa-X %100, spa-IgG %88.2 olarak tespit edilmiştir.
Spa-IgG 810 bç ve 970 bç, spa-X bölgenin 253 bç bir büyüklükte amplikon ürettiği
3. MATERYAL VE YÖNTEM
3.1. Materyal
Çalışma materyalini S.Ü. Veteriner Fakültesi, Mikrobiyoloji Laboratuarı’nda Emine ARSLAN tarafından tanımlanan Konya Bölgesi’ndeki mastitisli ineklerden izole edilen S. aureus izolatları oluşturmuştur.
3.1.1. Kimyasal maddeler ve besiyerleri
Fenol, kloroform, izoamil alkol gibi kimyasal maddeler Merck KGaA, 64271, Darmstadt, Almanya firmasından elde edilmiştir. Agaroz, etidyum bromür, NaCl, izopropanol, glasial asetik asit ise Sigma-Aldrich Chemie Gmbh, 89552, Steinheim, Besiyeri olarak Nutrient Broth (NB) ve Triptik Soy Agar (TSA) Oxoid marka kullanılmıştır.
3.1.2. Tampon ve çözeltiler
- TE tamponu : Tris 10mM, EDTA 1mM, pH=8 - Proteinaz K : 20mg/ml
- CTAB : %1 - SDS : %10 - NaCl : 5M
- Lizostafin : 2mg/ml
- Kloroform/İzoamil alkol : 24:1 oranında Kloroform/İzoamil alkol karıştırılarak hazazırlanmıştır.
- Fenol/Kloroform : 25:24 oranında - İzopropanol
3.2. Metod
3.2.1. ÇalıĢmada kullanılan bakteri kültürleri
-80°C`de saklanan bakteri suşları Triptik Soy Agar (TSA) besiyerinde aktif hale getirilmiştir. Daha sonra DNA izolasyonları için Nutrient Broth (NB) sıvı besiyerine ekim yapılarak 37°C`de 18 saat inkübasyona bırakılmıştır.
3.2.2. Bakteriden genomik DNA izolasyonu
TSA’ da aktif hale getirilen bakteriler, 5 ml’lik sıvı besiyerinde üretilerek 5000 rpm’de 1 dk santrifüj edilmiştir. Çökelti Tris-EDTA (TE) tamponunda pipet yardımıyla süspanse edilerek % 10 Sodyum dodesil sülfat (SDS) ve proteinaz K eklenip 37 o
C de 1 saat inkübe edilmiştir. 5 M NaCI eklenerek iyice karıştırılmıştır. Daha sonra Cetyl Trimethyl Ammonium Bromide CTAB/NaCI solüsyonu ilave edilerek iyice karıştırıldıktan sonra 10 dk 65 o
C’de inkübe edilmiştir. Eşit hacimde kloroform/ izoamilalkol ilave edilerek 5 dk 10,000 rpm’de santrifüj edilmiştir. Süpernatant’a eşit hacimde fenol/kloroform/izoamilalkol eklenip 10 dk santrifüj edilmiştir. Süpernatant yeni mikrosantrifüj tüpüne aktarılarak, izopropanol eklenerek ve 10 dk santrifüj yapılmıştır. Pellet %70’lik etanol ile yıkanıp kurutulduktan sonra 100 μl TE tamponunda bakteriyel DNA çözülerek -20’de saklanmıştır (Welsh ve McClelland, 1990). DNA konsantrasyonu spektrofotometre ile belirlenmiştir.
3.2.3. PCR uygulaması
Total hacim 30 μl olacak şekilde tüplere; 1 μl ileri primer (Forward), 1 μl geri primer (Reverse) (Çizelge 3,1.) 0,6 μl dNTP (dATP, dGTP, dCTP, dTTP), 3 μl 10x PCR tamponu, 1,8 μl MgCI2, 0,1 μl Taq polimeraz, 20 μl distile su ve son olarakta 2.5 μl DNA örneği ilave edilmiştir. Reaksiyon koşulları, denatürasyon bağlanma ve uzama aşamaları 30 döngü yapacak şekilde uygulanmıştır (94 o
C de 3 dk, 94 oC 1 dk, 51 oC 1 dk, 72 oC 1 dk, 72 oC 5 dk). Sıcaklıklar her bir primerin erime sıcaklığı (Tm) göz önüne alınarak yapılmıştır (Stephan ve ark., 2001).
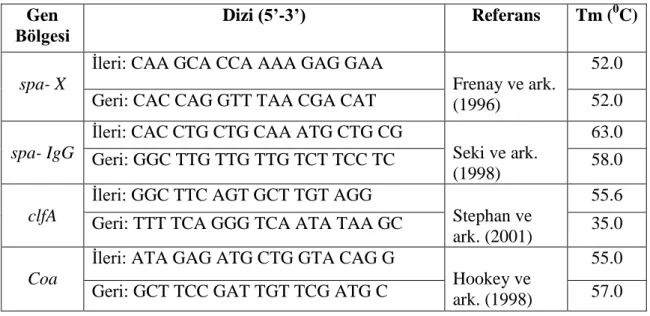
Çizelge 3.1. PCR uygulaması için kullanılan primerler Gen Bölgesi Dizi (5’-3’) Referans Tm (0C) spa- X
İleri: CAA GCA CCA AAA GAG GAA
Frenay ve ark. (1996)
52.0
Geri: CAC CAG GTT TAA CGA CAT 52.0
spa- IgG
İleri: CAC CTG CTG CAA ATG CTG CG
Seki ve ark. (1998)
63.0
Geri: GGC TTG TTG TTG TCT TCC TC 58.0
clfA
İleri: GGC TTC AGT GCT TGT AGG
Stephan ve ark. (2001)
55.6
Geri: TTT TCA GGG TCA ATA TAA GC 35.0
Coa
İleri: ATA GAG ATG CTG GTA CAG G
Hookey ve ark. (1998)
55.0
Geri: GCT TCC GAT TGT TCG ATG C 57.0
3.2.4. S. aureus suĢlarının PCR ürünlerinin agaroz jel elektroforezi
PCR ürünlerini değerlendirmek için %1.5’luk agaroz jel elektroforezine tabi tutulmuştur. Agaroz jel için 4.5 gr agaroz tartılıp, 300 ml TAE tamponu içinde kaynatılmıştır. Kaynadıktan sonra 15 μl etidyum bromür eklenerek elektroforez kabına dökülmüş ve taraklar yerleştirilmiştir. Daha sonra soğumaya bırakılmıştır. Jel soğuduktan sonra PCR ürünleri loding dye ile birlikte kuyucuklara yüklenerek 120 voltta yürütülerek U.V. transilluminatorde görüntülenen fotoğraf bilgisayara aktarılmıştır.
3.2.5. Veri analizi
Çalışılan genlere ait elde edilen sonuçlara göre spa-X, spa-IgG, clfA ve coa gen bölgeleri içeren örneklerin prevelansının %’lik değerleri hesaplanarak belirlenmiştir. Bu genlere ait büyüklükler ve oranlar saptanmıştır.
4. ARAġTIRMA SONUÇLARI VE TARTIġMA
4.1. S. aureus suĢunda spa-X, spa-IgG, clfA ve coa genlerinin PCR sonuçları
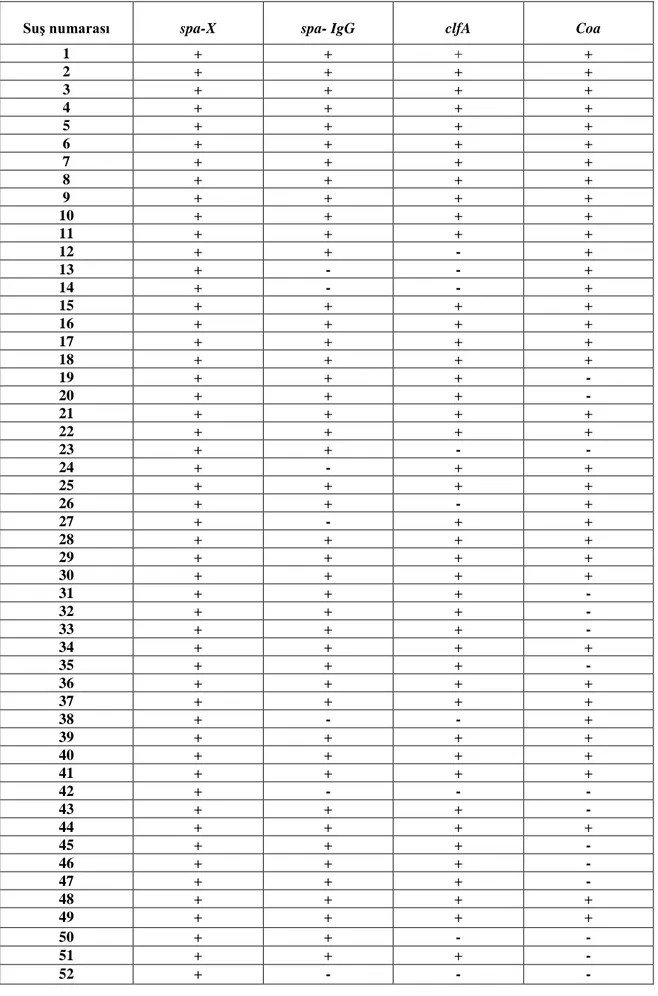
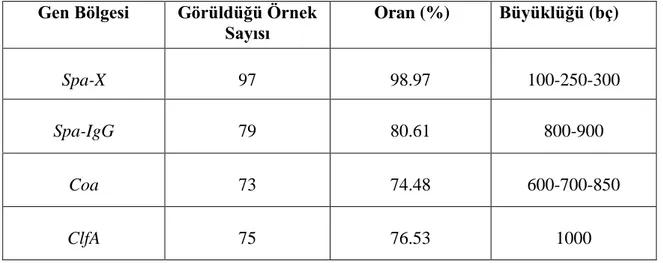
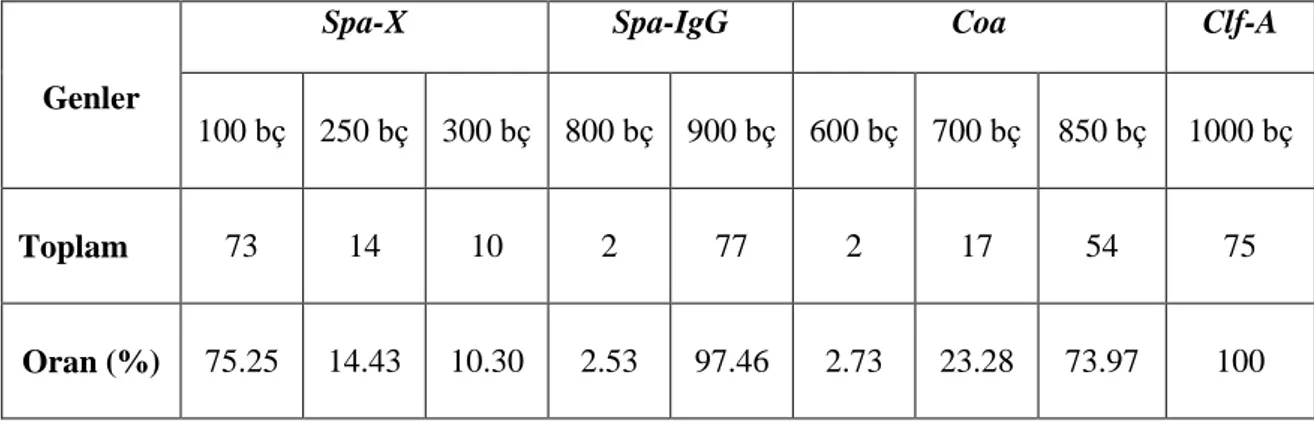
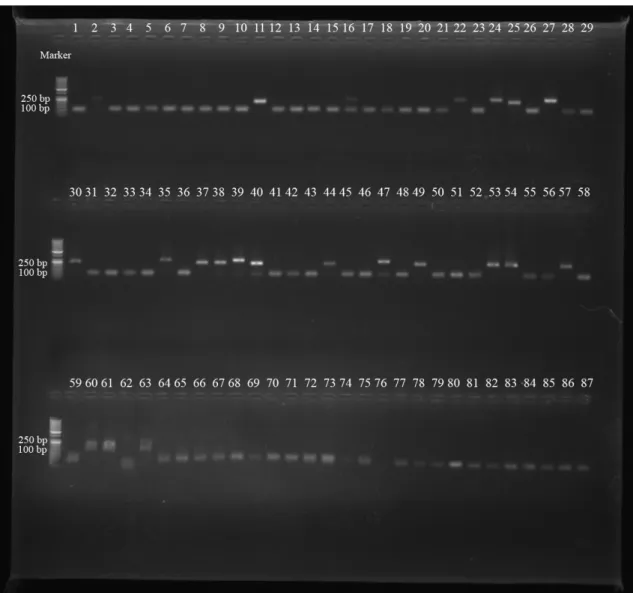
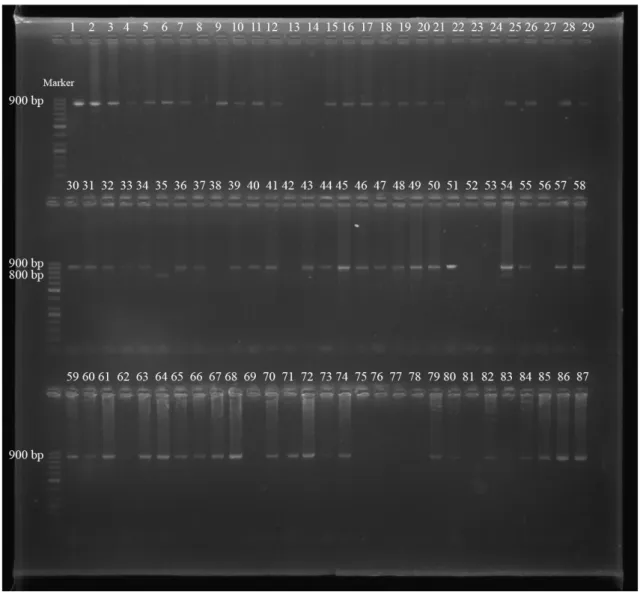
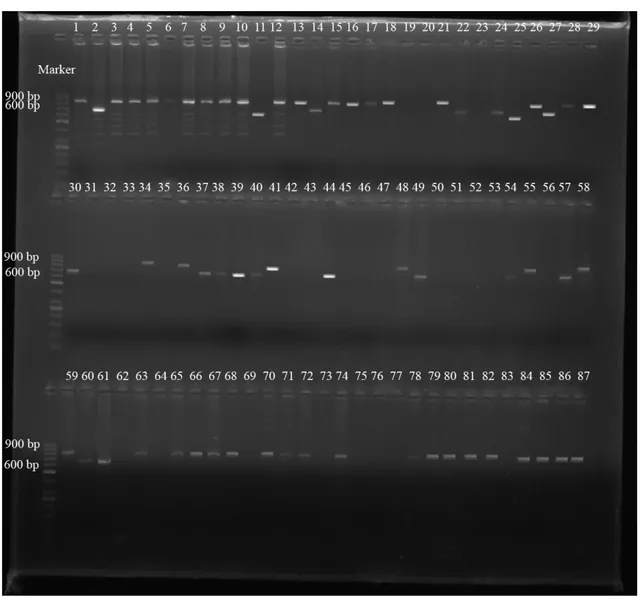
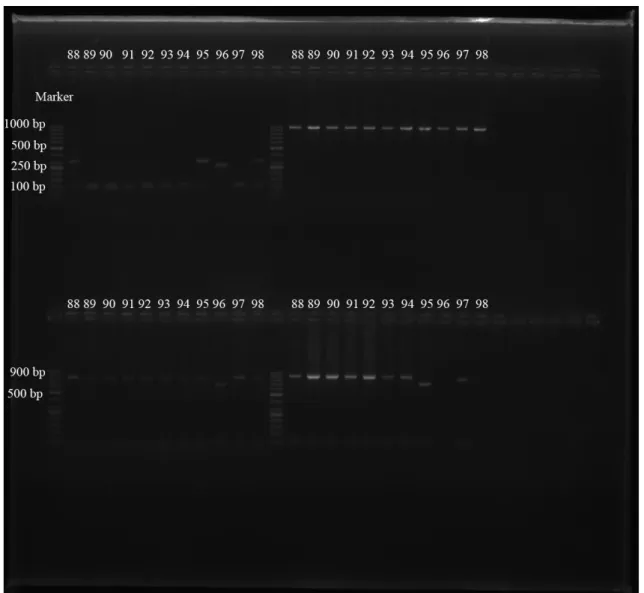
Çalışılan 98 S. aureus suşlarının spa-X, spa-IgG, clfA ve coa gen bölgeleri birer çift primer aracılığıyla çoğaltılmıştır. PCR ürünleri, agaroz jel elektroforezinde ethidium bromid ve DNA marker (50 bç) kullanılarak gözlenmiştir (Şekil 4.1., 4.2., 4.3., 4.4., 4.5.). 98 S. aureus suşunun 97’sinin spa-X geni (%98.97), 79’unun spa-IgG geni (%80.61), 75’inin clfA geni (%76.53), 73’ünün coa geni (%74.48) taşıdığı bulunmuştur (Çizelge 4.1., 4.2.).
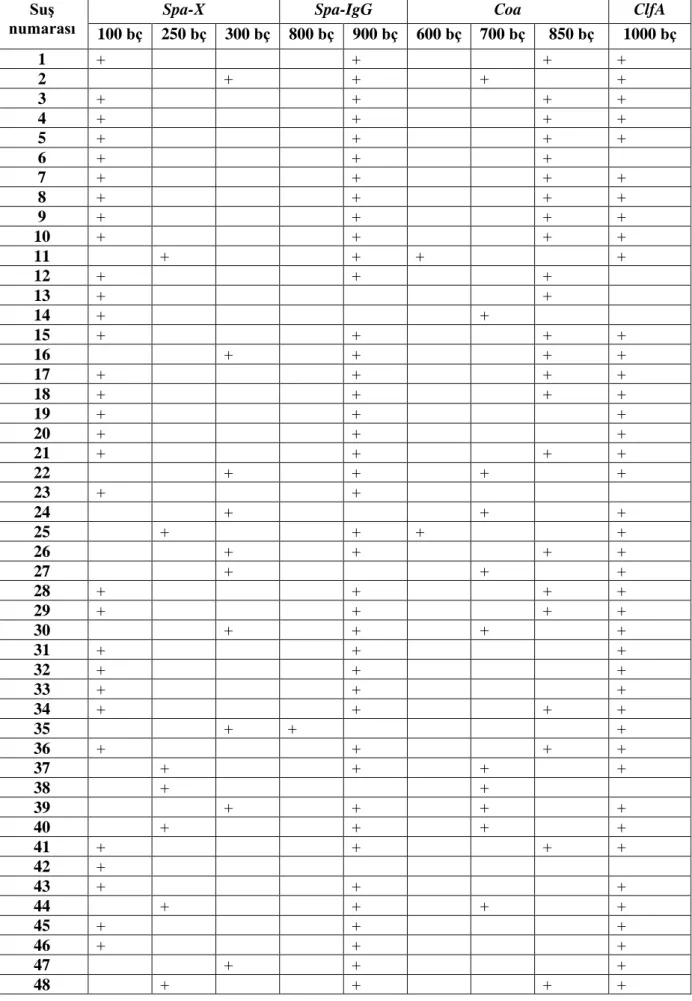
spa-X geninin 73’ünde (%75.25) 100 bç, 14’ünde (%14.43) 250 bç, 10’unda
(%10.30) 300 bç, spa-IgG geninde suşlardan 2’sinde (%2.53) 800 bç ve 77’sinde (%97.46) 900 bç, clfA geninde suşların 75’inde (%100) 1000 bç, coa geninde suşların 2’sinde (%2.73) 600 bç, 17’sinde (%23.28) 700 bç ve 54’ünde (%73.97) 850 bç büyüklükte bantlar elde edilmiştir (Çizelge 4.3., 4.4.).
Çizelge 4.1. S. aureus suşlarındaki gen bölgelerinin PCR sonuçları
SuĢ numarası spa-X spa- IgG clfA Coa
1 + + + + 2 + + + + 3 + + + + 4 + + + + 5 + + + + 6 + + + + 7 + + + + 8 + + + + 9 + + + + 10 + + + + 11 + + + + 12 + + - + 13 + - - + 14 + - - + 15 + + + + 16 + + + + 17 + + + + 18 + + + + 19 + + + - 20 + + + - 21 + + + + 22 + + + + 23 + + - - 24 + - + + 25 + + + + 26 + + - + 27 + - + + 28 + + + + 29 + + + + 30 + + + + 31 + + + - 32 + + + - 33 + + + - 34 + + + + 35 + + + - 36 + + + + 37 + + + + 38 + - - + 39 + + + + 40 + + + + 41 + + + + 42 + - - - 43 + + + - 44 + + + + 45 + + + - 46 + + + - 47 + + + - 48 + + + + 49 + + + + 50 + + - - 51 + + + - 52 + - - -
SuĢ numarası Spa-X Spa-IgG ClfA Coa 53 + - - - 54 + + + + 55 + + - + 56 + - - - 57 + + + + 58 + + + + 59 + + + + 60 + + + + 61 + + + + 62 - - - - 63 + + - + 64 + + + - 65 + + + + 66 + + + + 67 + + + + 68 + + + + 69 + - - - 70 + + + + 71 + + + + 72 + + + + 73 + + - - 74 + + + + 75 + - - - 76 - - - - 77 + - - - 78 + - + + 79 + + + + 80 + + + + 81 + - - + 82 + + + + 83 + - + - 84 + + + + 85 + + - + 86 + + + + 87 + + - + 88 + + + + 89 + + + + 90 + + + + 91 + + + + 92 + + + + 93 + + + + 94 + + + + 95 + + + + 96 + - + + 97 + + + + 98 + - + +
Çizelge 4.2. S. aureus suşlarındaki gen bölgelerinin oranları ve büyüklükleri Gen Bölgesi Görüldüğü Örnek
Sayısı Oran (%) Büyüklüğü (bç) Spa-X 97 98.97 100-250-300 Spa-IgG 79 80.61 800-900 Coa 73 74.48 600-700-850 ClfA 75 76.53 1000
Çizelge 4.3. S. aureus suşlarındaki gen bölgelerinin PCR sonuçları SuĢ
numarası
Spa-X Spa-IgG Coa ClfA
100 bç 250 bç 300 bç 800 bç 900 bç 600 bç 700 bç 850 bç 1000 bç 1 + + + + 2 + + + + 3 + + + + 4 + + + + 5 + + + + 6 + + + 7 + + + + 8 + + + + 9 + + + + 10 + + + + 11 + + + + 12 + + + 13 + + 14 + + 15 + + + + 16 + + + + 17 + + + + 18 + + + + 19 + + + 20 + + + 21 + + + + 22 + + + + 23 + + 24 + + + 25 + + + + 26 + + + + 27 + + + 28 + + + + 29 + + + + 30 + + + + 31 + + + 32 + + + 33 + + + 34 + + + + 35 + + + 36 + + + + 37 + + + + 38 + + 39 + + + + 40 + + + + 41 + + + + 42 + 43 + + + 44 + + + + 45 + + + 46 + + + 47 + + + 48 + + + +
SuĢ numarası
Spa-X Spa-IgG Coa ClfA
100 bç 250 bç 300 bç 800 bç 900 bç 600 bç 700 bç 850 bç 1000 bç 49 + + + + 50 + + 51 + + + 52 + 53 + 54 + + + + 55 + + + 56 + 57 + + + + 58 + + + + 59 + + + + 60 + + + + 61 + + + + 62 + 63 + + + 64 + + + 65 + + + + 66 + + + + 67 + + + + 68 + + + + 69 + 70 + + + + 71 + + + + 72 + + + + 73 + + 74 + + + + 75 + 76 77 + 78 + + + 79 + + + + 80 + + + + 81 + + 82 + + + + 83 + + 84 + + + + 85 + + + 86 + + + + 87 + + + 88 + + + 89 + + + + 90 + + + + 91 + + + + 92 + + + + 93 + + + + 94 + + + + 95 + + + + 96 + + + 97 + + + + 98 + + +
Çizelge 4.4. S. aureus suşlarındaki gen bölgelerinin büyüklükleri ve oranları
Genler
Spa-X Spa-IgG Coa Clf-A
100 bç 250 bç 300 bç 800 bç 900 bç 600 bç 700 bç 850 bç 1000 bç
Toplam 73 14 10 2 77 2 17 54 75
ġekil 4.1. S. aureus suşlarındaki spa-X gen bölgesinin PCR Ürünlerinin agaroz jel elektroforezi (Marker: 50 bç)
ġekil 4.2. S. aureus suşlarındaki spa-IgG gen bölgesinin PCR Ürünlerinin agaroz jel elektroforezi (Marker: 50 bç)
ġekil 4.3. S. aureus suşlarındaki clfA gen bölgesinin PCR Ürünlerinin agaroz jel elektroforezi (Marker: 50 bç)
ġekil 4.4. S. aureus suşlarındaki coa gen bölgesinin PCR Ürünlerinin agaroz jel elektroforezi (Marker: 50 bç)
ġekil 4.5. S. aureus suşlarındaki spa-X, clfA, coa, spa-IgG gen bölgelerinin PCR ürünlerinin sırasıyla agaroz jel elektroforezi (Marker: 50 bç)
Daha önce yapılan çalışmalarda clfA geninin S. aureus’da bulunma oranları farklılık gösterse de genellikle yüksek oranlarda bulunmuştur. Bu çalışmalardan bazılarında %100 oranında clfA geni bulunmuştur (Reinoso ve ark., 2008; Kalorey ve ark., 2007). Kumar ve ark. (2011) suklinik ve klinik mastitisli suşlarda sırasıyla %93.3, %87, Karahan ve ark. (2011) %91.3, Proietti ve ark. (2010) %82,4, Momtaz ve ark. (2010) %73.25, Klein ve ark. (2011) %50.6 oranlarla clfA geninin bulunduğunu tespit etmişlerdir. ClfA geninin yüksek prevelansa sahip olduğunu gözlemleyen bu çalışmaları bizim çalışmalarımız %76.53 oranıyla desteklemektedir. Bu sonuçların aksine Santos ve ark. (2014) çalıştıkları suşların %19.2 ‘sinde bu gene rastlamışlardır. Bant büyüklükleri ise bazı çalışmalarda; 900 bç ve 1000 bç olmak üzere iki farklı büyüklük bulunmuştur (Karahan ve ark., 2011; Reinoso ve ark., 2008; Kumar ve ark., 2011) Bunların dışında diğer çalışmalar ise bizim çalışmamızdaki gibi 1000 bç büyüklüğünde tek bir bant gözlemlediğimiz gibi 1042 bç, ve 680 bç büyüklüklerinden sadece birinin bulunduğunu tespit etmişlerdir (Proietti ve ark., 2010; Kalorey ve ark., 2007; Karahan ve ark., 2011).
Spa-X geni bazı çalışmalarda %100 oranında bulunmuştur (Karahan ve ark.,
2011; Akineden ve ark., 2001; Kumar ve ark., 2011; Proietti ve ark., 2010; Stephan ve ark., 2001). Khan ve ark. (2013) %98.5, Algammal ve ark. (2013) %92.8, Momtaz ve ark. (2010) %80.23, Kalorey ve ark. (2007) %76.47 gibi yüksek oranlarında tespit etmişlerdir. Bu oranlar bizim çalışmamızda bulduğumuz %98.97 sonucunu desteklemektedir. Spa-X geninde bu çalışmalarda farklı büyüklükte bantlar gözlenmiştir. Bazı araştırmacılar 253 bç, 320 bç, büyüklüklerinden yalnızca birini gözlemlemişlerdir (Proietti ve ark., 2010; Momtaz ve ark. 2010). Bu çalışmada 100, 250 ve 300 bç olmak üzere 3 farklı amplikon tespit edilmiştir. Bu sonuçları destekleyen birden fazla büyüklük gözlemleyen çalışmalar ise şu şekildedir; Karahan ve ark. (2011)
spa-X genini 110, 140, 170, 190, 220, 240, 270, 290 ve 320 bç. Akineden ve ark. (2001)
110, 140, 220, 240, 270, 290, 320 bç, El- Sayed ve ark. (2010) 320 bç, 150 bç ve 100 bç, Stephan ve ark. (2001) 300, 280, 200, 100 bç,. Khan ve ark. (2013) 190 ile 320 bç, Kalorey ve ark. (2007) 220, 253, 315 bç büyüklüklerinde, Reinoso ve ark., (2008) ise 100 bç dan 315 bç’e kadar oldukça polimorfik boyutlarda bulmuşlardır.
Spa-IgG geni bazı çalışmalarda %100 oranında bulunmuştur (Karahan ve ark.,
2011; Akineden ve ark., 2001; Kumar ve ark., 2011). Bizim çalışmamızda tespit ettiğimiz %80.61 oranına yakın çalışmalar ise; Kalorey ve ark. (2007) %91.89, Proietti ve ark. (2010) %88.2, gibi yüksek oranlarda bu gene rastlamışlardır. Bunların aksine Momtaz ve ark. (2010) %25.58 gibi düşük prevelansta Spa-IgG genini
gözlemlemişlerdir. Bazı araştırmacılar bizim çalışmamızda ulaştığımız 800 bç ve 900 bç olmak üzere iki büyüklükteki sonuçlara yakın olarak 700 bç ve 900, 920 bç ve 750 bç'lik iki farklı büyüklük tespit ederken (Coelho ve ark., 2008; Reinoso ve ark., 2008; Karahan ve ark., 2011; Stephan ve ark., 2001) bazıları 920 bç büyüklüğünde tek bir büyüklük gözlemlemişlerdir (Akineden ve ark., 2001; Momtaz ve ark., 2010). Proietti ve ark. (2010), 810 bç ve 970 bç, Kalorey ve ark. (2007) 590 bç, 810 bç ve 970 bç, Kumar ve ark. (2011) 500 bç, 1000 bç, 1050 bç olarak farklı boyutta bant tespit etmişlerdir.
Coa geni çalışmaların çoğunda %100 oranında bulunmuştur (Akineden ve ark.,
2001; Khan ve ark., 2013; Algammal ve ark., 2013; Kalorey ve ark., 2007). Kumar ve ark. (2011) % 93.3 oranıyla bizim çalışmamızda tespit ettiğimiz %74.48 oranı desteklerken Momtaz ve ark. (2010) %48.83 olarak daha düşük oranda gözlemlemişlerdir. 600 bç, 700 bç ve 850 bç büyüküklerinde üç farklı büyüklükte tespit ettiğimiz coa geni; Coelho ve ark. (2008) yaklaşık olarak 400 bç, 600 bç, 700 bç ve 900 bç, Akineden ve ark. (2001) 840 bç, 600 bç, 440 bç, Momtaz ve ark. (2010) 970 bç 730 bç, Stephan ve ark. (2001) 580-660 bç, Khan ve ark. (2013) 490 bç ile 204 bç; Kalorey ve ark. (2007) 627 bç, 710 bç, 910 bç büyüklüklerinde gözlemlemişlerdir. Daha önce yapılan bu çalışmalarda mastitisli sığırlardan izole edilmiş S. aureus’daki
clfA, coa, spa-X, spa-IgG virulans genlerinin yüksek oranda bulunması bizim
çalışmamızı desteklemektedir.
Spa’nın kagülaz ve nükleaz aktiviteleri ile büyük oranda korelasyon göstermesi, antifagositik etki yaratması ve antibiyotik duyarlılığını azaltması gibi özellikleri patojenite kriteri olacağına işaret etmektedir (Demirpek, 2006). Çoğu S. aureus suşlarının yüzeyi peptidoglikan ya da sitoplazmik membrana bağlanan protein A ile kaplanmıştır (Iraz, 2008). Bağlı koagülaz hücre duvarına bağlı halde fibrinojeni fibrine çevirerek hücre yüzeyinde fibrin presipitasyonu meydana getirir. Bunun sonucu olarak stafilokoklar aglütinasyon ve kümeleşmeye uğrar. Bakterinin patojenitesindeki rolü, fagositozu önlemek üzere bakterinin üzerine fibrin tabakası örtmesinden ileri gelmektedir (Boz, 2009). Klamping faktör A (clfA), S. aureus’un konakçı hücreye tutunması için en önemli adezyon kuvvet faktörlerinden biridir. ClfA, çözünebilir veya immobilize fibrinojen ile bakteriyel bağlanmayı kolaylaştırarak enfeksiyon sürecine katılmaktadır. Hayvan enfeksiyonlarında clfA’nın polimorf nüklear lökositlerin bakteriyi fagositozunu engelleme yeteneği önemlidir (Mcdevit ve ark., 19994; Karahan ve ark., 2005).
5. SONUÇLAR VE ÖNERĠLER
Çalışılan 98 S. aureus suşunun spa-X, spa-IgG, clfA ve coa gen bölgeleri birer çift primer aracılığıyla çoğaltılmıştır. PCR ürünleri agaroz jel elektroforezinde etidium bromide ve DNA marker kullanılarak gözlenmiştir (Şekil, 4.1., 4.2., 4.3., 4.4., 4.5.). 98
S. aureus suşunun 97’sinin (%98.97) spa-X geni, 79’unun (%80.61) spa-IgG geni,
75’inin (%76.53) clfA geni, 73’ünün (%74.48) coa geni taşıdığı bulunmuştur (Çizelge, 4.1).
spa-X geninde suşların 73’ünün (%75.25) 100 bç, 14’ünün (%14.43) 250 bç,
10’unun (%10.30) 300 bç, spa-IgG geninde suşların 2’sinin (%2.53) 800 bç ve 77’sinin (%97.46) 900 bç, clfA geninde suşların 75’inin (%100) 1000 bç, coa geninde suşların 2’sinin (%2.73) 600 bç, 17’sinin (%23.28) 700 bç ve 54’ünün (%73.97) 850 bç büyüklüğünde bantlar bulundurduğu gözlenmştir.
Virulans genler bakterinin patojenitesini arttırır. Virulans faktörler konakçının immun cevabından kaçması için önemlidir. Enfeksiyona neden olan bakterinin virulans genlerinin belirlenmesi ve tiplendirilmesi hastalığın tedavi sürecine katkı sağlayabilir. Bu tiplendirme, aşı çalışmaları için bakteri seçimini kolaylaştıracaktır. Bu sonuçlar mastitise karşı etkili koruma stratejilerini geliştirmede faydalı veriler sağlayarak tedavi sürecini kısaltacaktır. Bu sayede hayvanın erken mezbahaya gönderilmesi, süt verimi ve kalitesinin düşmesi gibi ekonomik kayıpların önüne geçilebilecektir.