DNA'nın arkeozomlarla formülasyonu, in vitro karakterizasyonu ve memeli hücre kültürlerinde taşıyıcı olarak kullanılması
Tam metin
Şekil




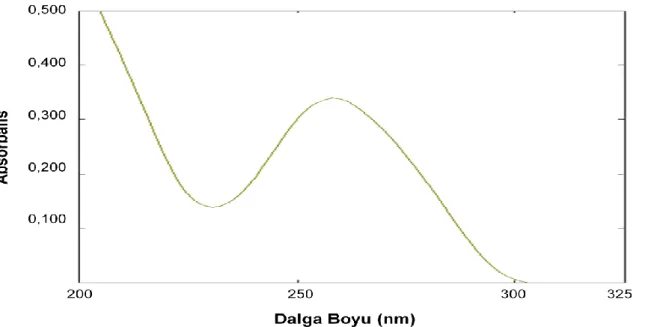
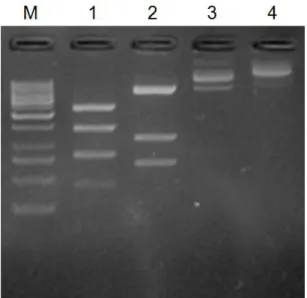
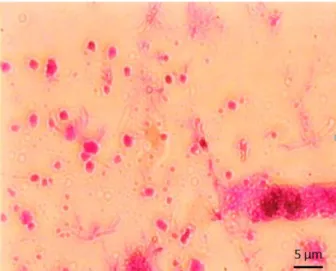
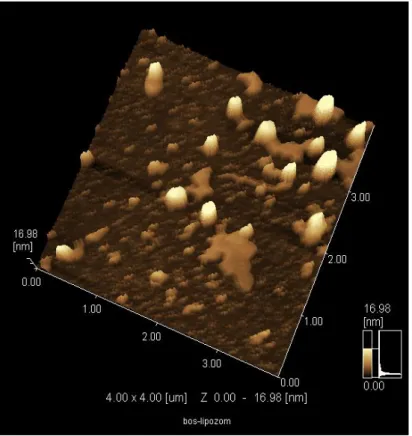
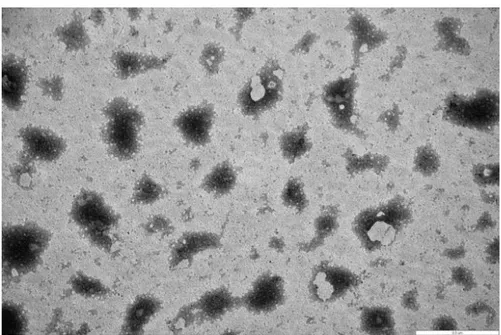

Benzer Belgeler
• Oositleri fertilize olabilir oosit aşaması olan Metafaz II aşamasına ulaşmasını sağlamak işlemidir. • In-vitro
Laboratuvar koşullarında embriyo üretimi anlamına gelir... In vitro maturasyon
• Fertilize the mature oocyte with capasitated semen to create an embryo. • In-vitro
1974 Smyth ve Davies Koyun ve at orijinli hidatik kistlerden elde edilen protoskolekslerin in vitro kültürüyle farklı E.granulosus fizyolojik suşlarının ortaya
PK/PD indeksleri, antibiyotiklerde in-vitro etkinliği in-vivo etki ile ilişkilendirerek, yeni antibiyotiklerin geliştirilmesi, kullanımda olan- ların doz ayarlamalarının
Noninvasive metabolomic profiling of embryo culture media using Raman and near-infrared spectroscopy correlates with reproductive potential of embryos in women undergoing in
Yapılan çalıĢmada bitkisel materyal olarak kullanılan koltuk altı tomurcuklarının BAP, IBA, GA 3 bitki büyüme düzenleyicileri kombinasyonları içeren besin ortamında
DSC analyzes of coarse DZ, stabilizers (Pluronic F127, PVP K30, and SDS), the lyophilized nanosuspension formulations (F-A and F-B) were performed at 25-400°C with a heating rate
