Karasal ortamlardan izole edilen streptomyces türlerinin sekonder metabolit biyosentez genlerinin taranması
Tam metin
Şekil
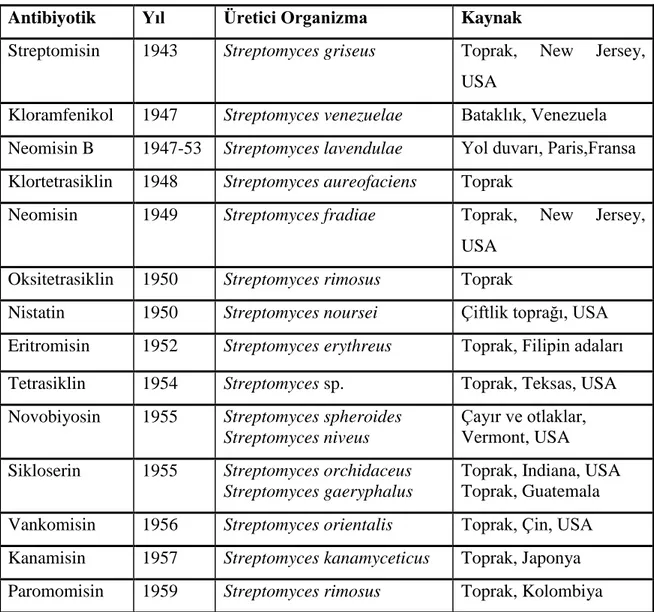
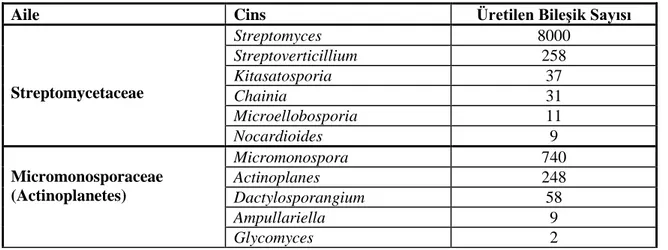

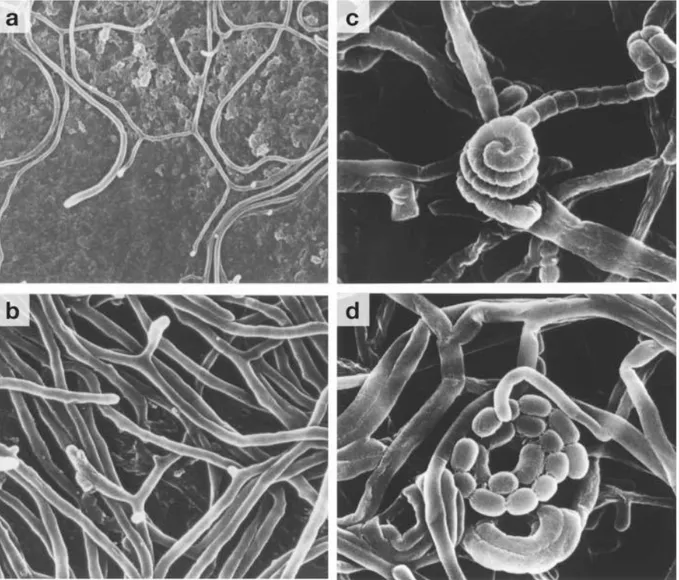
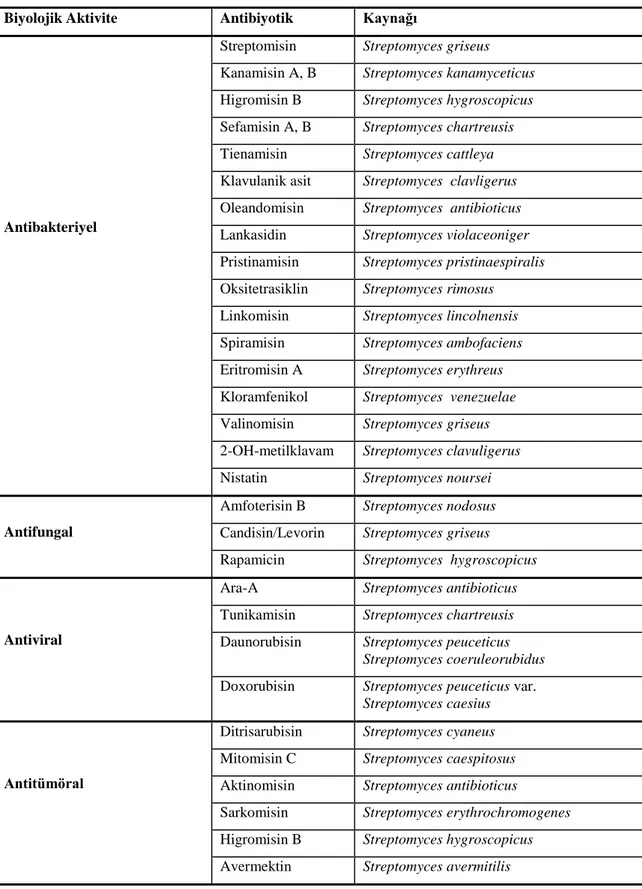
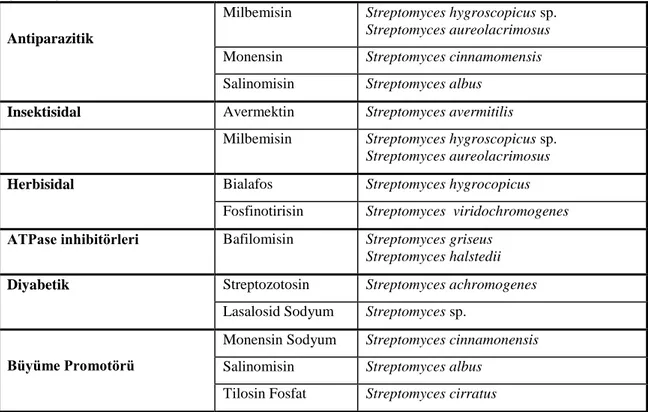
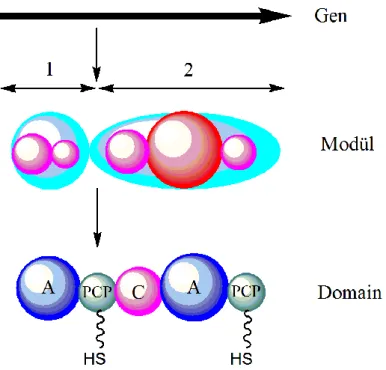

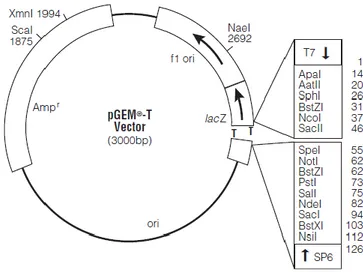

Benzer Belgeler
Aerobik ortamda ise, 3 ayrı talaş örneği (Carpinus betulus, Populus canadensis ve Pinus sylvester) ile hazırlanan mediumların inkübasyonu sonucu etil alkol üretip
In Kahramanmaras, Dwarf mistletoe (Arceuthobium oxycedri) was seen on Small fruited pine ( Juniperus oxycedrus subsp. oxycedrus ), Crimean juniper ( Juniperus
More recently some authors reported adipose tissue-derived stem cells and platelet- rich plasma leads to the regeneration of medullary bone-like tissue and long-term reduction of
Cerrahi öncesi görüntüleme olarak kranial kemik defekti için BT (aksiyal, kronal, sagital rekonstrüksiyonla), kese içeriği için MRG, (gliotik/fonksiyonel doku var mı?),
Toplumumuzda yayg›n olan mitlerin sönmesi, daha sa¤l›kl› ve do- yumlu bir cinsel yaflam için, cinsel bilgilerin zaman›nda ve do¤ru kaynaklardan bireylere aktar›l- mas›
Aynı değerlendirmeleri Şekil 4.8.’de KYME yakıt karışımları için yapacak olursak; KYME-20 yakıt için normal motorda birinci yük değeri altındaki termal verim
Etiology of uveitis % Anterior uveitis Idiopathic uveitis 31.5 Herpetic uveitis 15.5 Fuchs’ uveitis 11.6 Spondylarthropathies 11.1 HLA B27+ uveitis 10.9 Behçet’s disease 9.7
İnovasyonu kalite artışı sağlayacak bir süreç şeklinde algılayan yöne- ticilerin, inovasyon sürecini dönüşümcü tarzda yönetmeleri; İnovasyonu krizlerden çıkış olarak
