A new set of monoclonal antibodies directed to proline-rich and central regions of p53
Tam metin
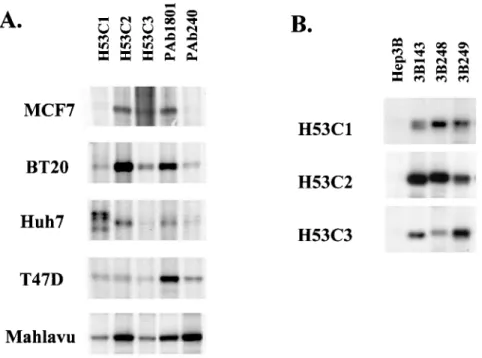
Şekil

Benzer Belgeler
İNSAN-I KÂMİL YAZI RESİMLERİNİN İKONOGRAFİK VE SEMBOLİK ANLAMLARINA DAİR BİR ÇÖZÜMLEME. Mürüvet
maddesine uyan suçu oluşturacağının gözetilmesinde zorunluluk bulun- ması, (Yarg. 6.CD., 22.2.2007, 8330/1721 sayılı kararı.) Hırsızlık suçundan yakalandığında
Rekabet hukuku yaptırımlarında tazminat davalarının en geniş şekilde uygulandığı ülke olma özelliğine sahip olan ABD, geliştirdiği sınıf davaları ile bu
Onun her şeyin hür nedeni olması, her şeyi irade hürlüğü ya da mutlak keyif ve hevesle değil, mutlak (zorunlu) tabiatının yani sonsuz gücünün eseri olarak
Hatta onun ismindeki çeşitlilik (söz gelişi Dinler Bilimi, Din Bilimi, Mukaye- seli Dinler Tarihi, Mukayeseli Din, Mukayese- li Dinler, Din Tarihi hatta Dinî Bilimler gibi)
Zakir Husain always realized the urgency of educational reforms and, therefore, deeply involved himself in evolving a scheme of national
Using this example as a guide, we define the integral
Even though government policy in less development countries as is the case with Cameroon acknowledges people with disabilities textually, application is still
