Aysegul Çebi, Egemen Akgün1, Serap Çelikler2, Mehmet Firat3, Mustafa Zafer Özel4, Engin Ulukaya5, Ferda Ari2 Department of Midwifery, Faculty of Health Sciences, 1Department of Medical Biology, Faculty of Medicine, Giresun University, Giresun, 2Department of Biology, Faculty of Science, Uludag University, Bursa, 3Department of Biology, Faculty of Education, Yuzuncu Yil University, Van,
5Department of Biochemistry, Faculty of Medicine, Istinye University, Istanbul, Turkey, 4Department of Chemistry, University of York, Heslington, York YO10 5DD, United Kingdom For correspondence: Dr. Aysegul Çebi, Faculty of Health Sciences, Giresun University, Piraziz, Giresun, Turkey. E‑mail: cebiaysegul@ hotmail.com
Cytotoxic and genotoxic effects of
an endemic plant of Turkey Salvia
kronenburgii on breast cancer cell lines
ABSTRACT
Context: The natural products derived from plants are the important sources that can be used for breast cancer treatment. Salvia species and their derived products were recommended as potential antitumor substances.
Aim: The potential cytotoxic and genotoxic effects of Salvia kronenburgii have been investigated on breast cancer cell lines, MCF‑7 and MDA‑MB‑231.
Materials and Methods: Determination of chemical compounds of S. kronenburgii was done using a gas chromatography coupled to time‑of‑flight mass spectrometry system and a dual‑stage commercial thermal desorption injector. Growth inhibition of the S. kronenburgii was measured by the 3‑(4,5‑dimethylthiazol‑2‑yl)‑2,5‑diphenyltetrazolium bromide and ATP viability assays. The cell death mode was detected by fluorescent dyes. Genotoxic effect of S. kronenburgii was measured by comet assay.
Results: S. kronenburgii showed antiproliferative effect in a dose‑dependent manner on MCF‑7 and MDA‑MB‑231 cell lines by inducing apoptosis‑like cell death. The pyknotic cell nuclei were observed at the cell lines in response to S. kronenburgii. Furthermore, significant increase was shown in genetic damage index and frequencies in the damaged cells.
Conclusion: S. kronenburgii might be a promising natural source for cancer therapy. Further experiments need to be done in vivo to understand of the anticancer effects of this plant.
KEY WORDS: Breast cancer, cytotoxicity, genotoxicity, in vitro, Salvia kronenburgii
Original Article
INTRODUCTION
Breast cancer is the most common malignancy among women worldwide. The American Cancer Society recently declared that 40,730 breast cancer
deaths have been estimated in 2015.[1] Thus, the
researches about anticancer active compounds from the plants have gained acceleration in the last decades. The genus Salvia L., with about 900 species, is one of the most widespread members of the family Lamiaceae.[2] It is represented in Turkey by 94 taxa
belonging to 89 species with 50% endemism.[3] The
Lamiaceae is the wide range of plant family which is known to have antioxidant capacity due to phenolic
compounds.[4] In a study, eight Salvia species were
investigated, and all of these had high antioxidant
activity, in Turkey.[5] Salvia kronenburgii is a Turkish
endemic plant, and there are not enough data about cytotoxic effect of this endemic species.
A few Salvia species are known to cure some diseases such as cancer, microbial infections,
and malaria for local traditional uses.[6] It
has been demonstrated that some Anatolian Salvia species have cytotoxic activity against
the human ovarian cancer cell line.[7] DNA
fragmentation and reactive oxygen species induction are associated with anticancer effect
of CH2Cl2 fraction obtained from Symplocos
chloroleuca.[8]
The new endemic plants come into prominence at this scientific ambient. The new active compounds obtained from some Salvia species are leading promising researches for the anticancer drug design and development. Tanshinone, salvicine, and their derived compounds have been researched
as anticancer drugs.[9‑11]
Access this article online Website: www.cancerjournal.net DOI: 10.4103/jcrt.JCRT_1322_16 Quick Response Code:
This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution‑NonCommercial‑ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non‑commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
For reprints contact: [email protected]
Cite this article as: Çebi A, Akgün E, Çelikler S, Firat M, Özel MZ, Ulukaya E, et al. Cytotoxic and genotoxic effects of an endemic plant of Turkey Salvia kronenburgii on breast cancer cell lines. J Can Res Ther 2019;15:1080-6.
In the last years, cytotoxic and genotoxic activities of the extracts of some Salvia species have been investigated. Jantová et al. researched that the molecular mechanism of Salvia officinalis extract induced apoptosis on the L1210 leukemia
cells.[12] Russo et al.[13] investigated the growth inhibitory
and proapoptotic effect of 18 S. officinalis essential oils in
the human melanoma cell lines. Zhao et al.[14] reported that
polyphenol‑rich extract of Salvia chinensis showed anticancer activity in different cancer cell lines and induced apoptosis in pancreatic cancer cells.
This study was purposed to investigate the cytotoxic and genotoxic activities of an endemic plant S. kronenburgii on MCF‑7 and MDA‑MB‑231 human breast cancer cell lines. The volatile components of the S. kronenburgii extract were also determined by Gas chromatography coupled to time‑of‑flight mass spectrometry (GCXGC‑TOF/MS). To the best of our knowledge, this is the first report on the genotoxic activity of S. kronenburgii collected from Turkey.
MATERIALS AND METHODS
Collections and identification of plant materials
S. kronenburgii was collected from Gürpinar district, Kurubaş pass, Van, Turkey, in June 2010 and identified aid of flora
books.[15] The voucher specimens have been deposited at
the Herbarium of the Department of Biology of Yuzuncu Yil University, Van, Turkey.
Extraction of plant samples
The plant samples were air‑dried at room temperature, and then extraneous materials were carefully removed from the plants. The whole material (the trunk, flower, and root of samples) was got into powder, and 15 g of the plant material was extracted by adding 150 ml of solvent methanol (Merck, NJ, USA) in Soxhlet apparatus for 24 h. The crude extracts were concentrated using rotary evaporator at 40°C, and then the residues were lyophilized and stored at −20°C until utilized for the other tests.
Determination of chemical compounds of Salvia kronenburgii
Direct thermal desorption
The content of the trunk and flower parts of S. kronenburgii was determined by GCXGC‑TOF/MS method in York University, England. A GCXGC‑TOF/MS system and a dual‑stage commercial thermal desorption injector were used together. To eliminate the risk of contamination, about 20–30 mg of the sample was placed by tweezers into the thermodesorption tubes. Initial desorption of the sample was carried out by heating the thermal desorption unit from 40°C (initial time 0.2 min) to 150°C at a rate of 120°C/min with a final hold time of 10 min under a helium flow of 1.5 mL/min in splitless mode. Volatile analytes released from this heating were cryo‑focused at −40°C in the cooled injector system (CIS) cooled with liquid nitrogen before injection. The CIS was then heated at a rate of 10°C/s to a final temperature of 150°C. Analytes
were transferred splitless to the GC column during the CIS temperature ramp.
Chromatographic analysis
The GCXGC‑TOF/MS system consisted of an Agilent 6890 (Agilent Technologies, Palo Alto, CA, USA) gas chromatograph and a Pegasus III TOF‑MS (LECO, St. Joseph, MI, USA). The modulator between first and second GC columns was based on a Leco (Cheshire, UK) liquid nitrogen two‑stage cold jet system. The modulation time was 5 s. The first column was a nonpolar BPX5 (30 m × 0.32 mm i.d. × 0.25 μm film thickness) and the second column a BPX50 (1.5 m × 0.10 mm i.d. × 0.10 μm film thickness) both from SGE Analytical Science (VIC, Australia). The initial temperature of the first dimension column was 60°C for 1 min, and the subsequent temperature program was a heating rate of 5°C/min until 310°C was reached and held isothermally for a further 1 min. The initial temperature of the second dimension column was 75°C for 1 min, and a 5°C/min heating rate was used until 325°C was reached and held isothermally for further 1 min. Helium was used as a carrier gas at a constant flow of 1.0 mL/min. The first‑dimensional separation is based on the separation by volatility in a nonpolar column. The second‑dimensional separation is based on the separation by polarity using a more polar column. The combination of separations produces the overall two‑dimensional chromatogram. Peak identification was made using TOF/MS with electron ionization.
Determination of cytotoxicity
Chemicals and cell culture
Breast cancer cell lines (MCF‑7 and MDA‑MB‑231) were cultured in RPMI 1640 supplemented with penicillin G (100 U/ml), streptomycin (100 μg/ml), L‑glutamine, and 10% fetal calf serum (Invitrogen, Paisley, UK) at 37°C in a
humidified atmosphere containing 5% CO2. S. kronenburgii
was dissolved in dimethyl sulfoxide (DMSO) at 100 mg/ml concentration as a stock solution. Further dilutions were made in culture medium. S. kronenburgii extracts were used at different concentrations ranging from 0.05 to 100 μg/ml. The 3‑(4,5‑dimethylthiazol‑2‑yl)‑2,5‑diphenyltetrazolium bromide viability assay
The cells first cultured in 96‑well microplates (5 × 103/well in 200 μl medium) and then incubated with different concentrations of S. kronenburgii in medium for 48 h. The untreated cells received only the medium and solvent (0.1% DMSO as final concentration). Each experiment was carried out twice in duplicates. A 3‑(4,5‑dimethylthiazol‑2‑yl)‑2,5‑diphenyltetrazolium bromide (MTT) viability assay was performed as previously
described.[16] MTT was first prepared as a stock solution of
5 mg/ml in phosphate buffer (PBS, pH 7.2) and filtered. At the end of the treatment period (48 h), 20 μl of MTT solution (5 mg/ml PBS, pH 7.2) was added to each well. After overnight incubation, the absorbance was read by an ELISA plate reader (FLASH Scan S12, Analytik Jena, Germany) at 570 nm to determine the cell viability. The % viability of treated cells was
calculated by dividing the absorbance of treated cells with the absorbance of control cells and then by multiplying with 100. The ATP viability assay
The ATP assay, a highly sensitive luciferin: luciferase‑based assay, was performed to determine the level of cellular ATP
as an indirect marker for the number of alive cells.[17] This
assay was performed to confirm the results of the MTT assay as the ATP assay is more reliable and sensitive than
the MTT assay.[16] The cells were seeded, grown and treated
as explained in the MTT assay (see above). At the end of the treatment period (48 h), cell viability was determined by the ATP assay (ATP Bioluminescent Somatic Cell Assay Kit, Sigma, St. Louis, MO, USA) according to the standard protocol of the manufacturer.
Fluorescence imaging for determination of cell death mode
Determination of cell death mode was examined staining by fluorescent dyes, Hoechst 33342, Calcein‑AM, and propidium iodide (PI). Hoechst 33342 can bind DNA and stains living or dead cells as blue. Apoptotic cells are observed brighter and smaller when they stained with Hoechst 33342 dye. Calcein‑AM dye, a molecule which can pass the cell membrane, stains only the living cells as green. PI stains just dead cells as red owing to pass through damaged membranes. Thus, PI describes specifically the late apoptotic or secondary necrotic and primary necrotic cells. For this reason, MCF‑7 and MDA‑MB‑231 cells were seeded in a 96‑well plate
at the density of 1 × 104 cells per well in 100 μl culture
medium. Then, the cells were treated with S. kronenburgii extract for 48 h. Following the treatment, the cells were incubated for 20 min in the dark at 37°C with the staining solution (Hoechst 33342, PI, and Calcein‑AM) of Cell Viability Imaging Kit (Roche, Mannheim, Germany) as instructed by the manufacturer. Then, the cells were analyzed with a fluorescence microscope.
Determination of genotoxicity by comet assay
The alkaline comet assay was performed by some
modifications.[18] First, the breast cancer cells (MCF‑7 and
MDA‑MB‑231, 5 × 105 cells/well) were seeded in six‑well plates.
The cells were incubated with 100 μg/ml of S. kronenburgii extract. Ethyl methanesulfonate (EMS) was added to the wells as positive controls for 24 h. Trypsin buffer was added to the cells and suspended in PBS (60 μl) buffer. The cell
suspension (3 × 104 cells) was mixed with 1% low melting
point agarose and quickly laid on slides coated with 1% normal melting point agarose. The treated cells were lysed and then neutralized. Ethidium bromide (2 μg/ml) was used to stain the slides and visualized under a fluorescence microscope. The specialized software was used to analyze the images (Argenit, Istanbul, Turkey). The comet length, tail length, tail DNA%, olive tail moment, head DNA%, genetic damage index (GDI), and percentage of damaged cell parameters were measured for the cells. GDI was evaluated by the following formula used
by comet analysis program.[19]
GDI = Type1 Type2 Type3 Type4 Type0 Type ( ( ) ( ) ( ) ( 1 2 3 4 × + × + × + × +
∑
∑
∑
∑
∑
11 Type2 Type3 Type4∑
∑
∑
∑
+ + + )% Damaged cell =
∑
Type2+∑
Type3+∑
Type3Statistical analyses
All statistical analyses were performed using theIBM SPSS
ver. 20.0 (IBM Co., Armonk, NY, USA) for Windows. The significance was calculated using one‑way analysis of variance and Tamhane’s T2 tests. P < 0.05 was considered statistically significant. Results were expressed as mean ± standard error.
RESULTS
Chemical analyses of Saliva kronenburgii
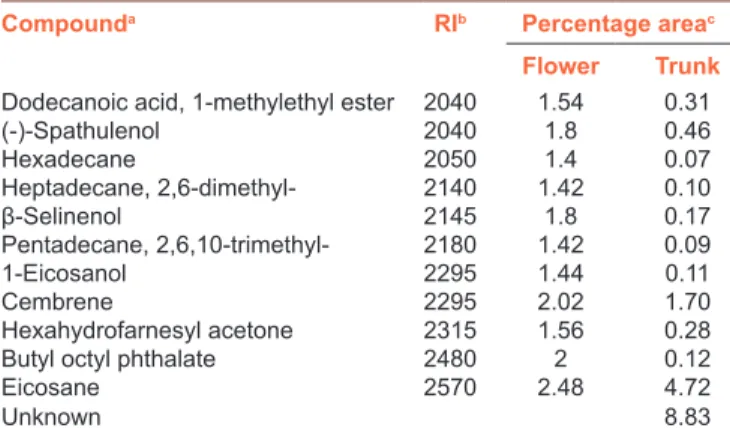
The chemical composition of S. kronenburgii was analyzed by GCXGC‑TOF/MS system and the qualitative and quantitative compositions are presented in Table 1. Sixty‑nine compounds were identified in S. kronenburgii and the major components of it were eucalyptol (27.15%), linalool oxide (11.94%), 2,2,4‑trimethylpentanediol‑1,3‑diisobutyrate (10.5%), acetic acid (5.73%), eicosane (4.72%), camphor ( 3 . 6 1 % ) , 1 , 2 ‑ c y c l o h e x a n e d i o l , 1 ‑ m e t h y l ‑ 4 ‑ (1‑methylethenyl)‑(2.85%), D‑carvone (2.25%), limonene (1.93%), trans‑carveol (1.88%), verbenone,(l) (1.82%), α‑terpineol (1.73%), cembrene (1.70%), and epoxylinalol (1.13%). All other components were present in <1%.
Antigrowth activity of Salvia kronenburgii
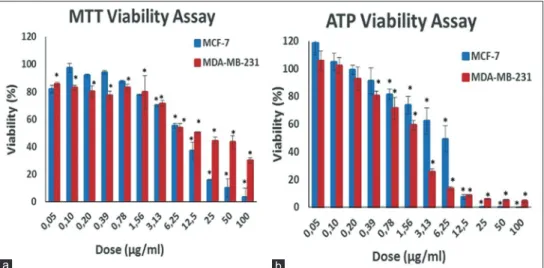
Antigrowth activity of S. kronenburgii (0.05–100 μg/ml, 48 h) was assessed on breast cancer cell lines (MCF‑7 and MDA‑MB‑231) by the MTT and ATP viability assays [Figure 1]. S. kronenburgii extract significantly inhibited the growth of the MCF‑7 and MDA‑MB‑231 cells in a dose‑dependent manner (P < 0.05). Subsequently, the results were confirmed by a more sensitive ATP viability assay [Figure 1b]. The ATP level significantly decreased after S. kronenburgii treatments at dose‑dependent in both types of cells (P < 0.05) [Figure 1b]. Half‑maximal inhibitory concentration (IC50) and 90% inhibitory concentration (IC90) values of S. kronenburgii were calculated on the basis of MTT and ATP assay results [Table 2]. According to the ATP assay results, IC50 values for S. kronenburgii were calculated 1.9 μg/ml and 6.0 μg/ml for MDA‑MB‑231 and MCF‑7 cell lines, respectively.
Fluorescence imaging for determination of cell death mode
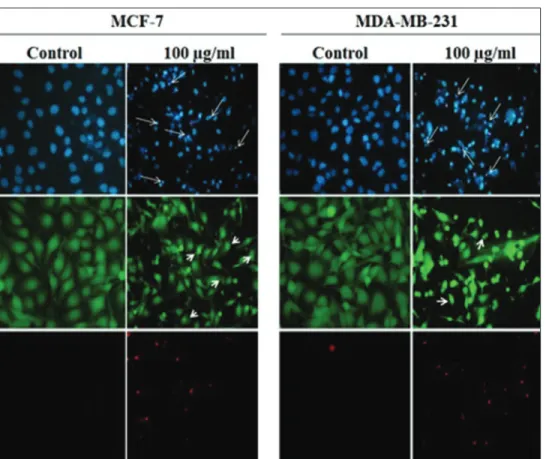
MTT and ATP assay results showed that S. kronenburgii displayed a potent cytotoxic activity at 100 μg/ml dose against the MCF‑7 and MDA‑MB‑231 cell lines. The cell death mode induced by S. kronenburgii was evaluated by fluorescence imaging on the basis of nuclear morphology. Hoechst 33342 dye can bind DNA and pass through the cell and stains all of the apoptotic or necrotic
cells. In this study, nuclei of the cells, which were stained with Hoechst 33342 dye, was observed smaller and brighter compared to control [Figure 2]. Calcein‑AM can pass through cell membrane, but it is not a fluorescent molecule. In living cells, esterases (a group of enzymes) interact with Calcein‑AM to produce calcein (fluorescent molecule). Only living cells can be stained by Calcein‑AM after treatment. The cells staining with Calcein‑AM were observed as green. On the other hand, the cells in the same field were not stained with PI dye (because of the cell membrane is intact, the cells are not observed as red) which is a characteristic feature of early apoptotic cell death. PI is a fluorescent dye that can stain all of the dead cells (late apoptotic/necrotic) and it effluxes by living cell. Therefore, it can only pass through cells that have membrane damage. To evaluate cell death mode, MCF‑7 and MDA‑MB‑231 cells were treated with S. kronenburgii (100 μg/ml) for 48 h. According to the results, we observed that S. kronenburgii resulted in cell shrinkage, pyknotic and fragmented nuclei in cells, which are well‑known feature of apoptosis [Figure 2].
Comet assay
The comet assay results of S. kronenburgii extract, positive control (EMS) and negative controls (at 0 h) in breast cancer cell lines are shown in Table 3. GDI values were significantly different from negative and positive controls at 4, 8, and 24 h for both cell lines (P < 0.0001). Damage frequencies were significantly different at 8 and 24 h for MCF‑7 whereas at 24 h for MDA‑MB‑231 (P < 0.0001). Tail length was statistically significant at 24 h for both cell lines (P < 0.0001 for MCF‑7, P < 0.05 for MDA‑MB‑231). Average comet density was significantly different at 4, 8, and 24 h in MCF‑7 cells (P < 0.0001). It was different at 2 and 24 h in MDA‑MB‑231 cells (P < 0.005, P < 0.0001, respectively). The percentage of tail DNA was statistically significant at 24 h in both of them [P < 0.0001, Table 3].
Table 1: Contd...
Compounda RIb Percentage areac
Flower Trunk
Dodecanoic acid, 1-methylethyl ester 2040 1.54 0.31
(-)-Spathulenol 2040 1.8 0.46 Hexadecane 2050 1.4 0.07 Heptadecane, 2,6-dimethyl- 2140 1.42 0.10 β‑Selinenol 2145 1.8 0.17 Pentadecane, 2,6,10-trimethyl- 2180 1.42 0.09 1-Eicosanol 2295 1.44 0.11 Cembrene 2295 2.02 1.70 Hexahydrofarnesyl acetone 2315 1.56 0.28
Butyl octyl phthalate 2480 2 0.12
Eicosane 2570 2.48 4.72
Unknown 8.83
aAs identified by GCXGC‑TOF/MS software; names according to NIST mass spectral library, and by comparing their Kovats retention indices, bKovats retention index of each component was collected from the literature for column Rxi 5ms, cPercentage of each component is calculated as peak area of analyze divided by peak area of total ion chromatogram times 100 (In the case of multiple identification, the areas of the peaks that belong to one analyze were combined to find the total area for this particular analyze). RI=Retention Index, NIST=National Institute of Standards and Technology, GCXGC-TOF/ MS=Gas chromatography coupled to time‑of‑flight mass spectrometry
Table 1: Total bioactive compounds of Salvia kronenburgii
Compounda RIb Percentage areac
Flower Trunk 3-Buten-2-ol, 2-methyl- 500 1.4 0.10 Formic acid 580 1.28 0.17 Acetic acid 610 1.38 5.73 Acetol 645 1.46 0.18 Propanoic acid 685 1.36 0.13 Allyl acetone 690 1.46 0.05 Furfural 810 1.72 0.09 1-Methoxy-2-propyl acetate 825 1.52 0.45 Nonane 865 1.32 0.05 Benzaldehyde 885 2.14 0.12 Carveol 960 1.64 0.27 α‑pinene 965 1.7 0.54 Phenol 985 1.96 0.05 Camphene 990 1.44 0.11 2,4-Thujadiene 995 1.48 0.32 2(5H)-Furanone, 5,5-dimethyl- 1020 1.88 0.42 (E)-3-Caren-2-ol 1030 1.68 0.22 2,3-Epoxycarane, (E)- 1035 1.66 0.21 β‑Pinene 1050 1.7 0.06 o-Cymene 1125 1.54 0.96 Terpinyl acetate 1130 1.48 0.38 Limonene 1145 1.62 1.93 Eucalyptol 1150 1.78 27.15 Linalool oxide 1150 1.68 11.94 Acetophenone 1170 2.02 0.09 Methylpyrrolidone 1220 2 0.08 m-Cresol 1250 1.8 0.10 Linalool 1255 1.54 0.16 Trans-2-Nonenol 1260 1.52 0.09 6-Methyl-3,5-heptadiene-2-one 1280 1.7 0.25 Cyclopropyl carbinol 1315 2.18 0.04 Camphore 1345 1.84 3.61 (E)-3(10)-Caren-4-ol 1360 1.7 0.29 1-Cyclohexanol, 1-vinyl- 1365 2 0.20 Pyranone 1385 2.04 0.93 Epoxylinalol 1405 1.68 1.13 Isoborneol 1415 1.7 0.80 p-Cymen-8-ol 1435 1.78 0.41 p-Acetotoluene 1440 1.88 0.34 α‑Terpineol 1445 1.66 1.73 Dihydrocarvone 1450 1.72 0.25 Verbenone,(l) 1485 1.86 1.82 Trans-Carveol 1485 1.76 1.88 Bergamiol 1495 1.54 0.18 Linalyl anthranilate 1500 1.54 0.10 cis-Carveol 1510 1.72 0.17 D-Carvone 1520 1.82 2.25 6-Methylheptane-1,6-diol 1525 2.12 0.08 2-Hexanol, 2,5-dimethyl-, (S)- 1535 2.14 0.11 1-Propanol, 3-phenoxy- 1535 1.92 0.14 1,2-Cyclohexanediol, 1-methyl-4-(1-methylethenyl)- 1710 1.94 2.85 Propanoic acid, 2-methyl-,
3-hydroxy-2,4,4-trimethylpentyl ester 1720 1.66 0.32 2-Dodecanone 1730 1.56 0.10 Cyclopropanecarboxylic acid, 1-(phenylmethyl)-, 2,6-bis (1,1-dimethylethyl)-4-methylphenyl ester 1865 1.62 0.13 Pentanoic acid, 2,2,4-trimethyl-3-carboxyisopropyl, isobutyl ester 1950 1.64 0.18 Leucoglucosan 1980 2.44 0.09 2,2,4-Trimethylpentanediol-1, 3-diisobutyrate 2010 1.74 10.50 Contd...
DISCUSSION
Herbal products have been commonly used for cancer therapies since ancient times. Breast cancer is the most common malignancy of woman worldwide, with more than one million new cases occurring annually. The discovery of the novel compounds is required to cure the breast cancer. Salvia species are known to have antioxidant effects as
they include high phenolic contents.[5] We analyzed the
chemical composition of this endemic plant, S. kronenburgii, using GCXGC‑TOF/MS system. Sixty‑nine compounds were found in this plant extract. Eucalyptol, linalool oxide, and 2,2,4‑trimethyl pentanediol‑1,3‑diisobutyrate were the most
common compounds in this plant. Oztekin et al.[20] isolated two
bioactive diterpenoids, horminone, and 7‑O‑acetylhorminone by using a micellar electrokinetic chromatography method for the simultaneous quantitative analysis in S. kronenburgii. In this study, we evaluated possible cytotoxic and genotoxic effects of S. kronenburgii extract on the breast cancer cell lines. Cytotoxic effect of the S. kronenburgii
(0.05–100 μg/ml, 48 h) was assessed on breast cancer cell
lines (MCF‑7 and MDA‑MB‑231) by the MTT and ATP assays. IC50 and IC90 values for S. kronenburgii are shown in Table 2. We found that S. kronenburgii extract had more cytotoxic
activity in MDA‑MB‑231 than MCF‑7 cell lines for ATP assay. In a study, growth inhibitory effect of Salvia officinalis was measured on human melanoma cell lines by MTT test. Essential oils of S. officinalis showed major cytotoxic effect on these
melanoma cell lines.[13]
Some studies have shown that Salvia species potential antitumor
effects for different tumor cell lines.[13] Salvia extracts have been
suggested to use preventing cancer such as melanoma and
leukemia.[13,21] Since the extracts of new endemic species provide
possibilities in vitro remarkable pilot studies, Ethanol extracts of two different Salvia species, Salvia triloba and Salvia dominica, were investigated for their antiproliferative activity against breast cancer cell lines (MCF7 and T47D) by the sulforhodamine
B assay. IC50 values of S. triloba and S. dominica were found
as 29.89 ± 0.92 and 38.91 ± 2.44 μg/mL while 5.83 ± 0.51
and 12.83 ± 0.64 μg/mL for against MCF7 and T47D cells,
respectively. The ethanolic extracts of these Salvia species were suggested as useful substances in breast cancer treatment via
proapoptotic cytotoxic mechanisms.[22] The methanolic extract
of S. officinalis was offered an inhibition of cell proliferation
by dose‑dependent manner in human leukemic cell lines.[23] In
another study, CH2Cl2 fraction obtained from S. chloroleuca was
defined as a potential apoptotic agent in MCF‑7 cell lines.[8]
The comet assay is used to distinguish apoptosis and necrosis
based on the DNA fragmentation pattern.[18] High values
of tail moments and distance between head and tail of the
comet can be related to apoptosis.[24] The tail length and
density also show the extent of strand breaks in DNA. Thus, a quantitative measure of the damaged DNA is performed by
the using the percentage of DNA in the tail.[25] In our study,
we evaluated the genotoxic effect of S. kronenburgii using the comet assay in MCF‑7 and MDA‑MB‑231 breast cancer cell lines for the first time. GDI values have been found as significantly different comparing with positive and negative controls by time depending manner (at 4, 8, and 24 h) for Table 2: Antigrowth parameters for Salvia kronenburgii
extract were determined by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide and adenosine
triphosphate assay after the treatment for 48 h
Cell line MTT assay ATP assay
IC50
(μg/ml)* (μg/ml)IC90 (μg/ml)*IC50 (μg/ml)IC90
MCF-7 8.07 49.76 6.06 12.12
MDA-MB-231 12.95 175.75 1.99 10.57
*IC50 is defined as the dose inhibiting 50% of viability, *IC90 is defined as the dose inhibiting 90% of viability. ATP=Adenosine triphosphate, MTT=3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
Figure 1: Assessment of viability of MCF-7 and MDA-MB-231 cell lines after 48 h treatment with varying doses of Saliva kronenburgii extract by MTT (a) and ATP (b) viability assays.*Denotes statically significant differences in comparison with control (P < 0.05). MTT = 3‑(4,5‑dimethylthiazol‑2‑yl)‑2,5‑diphenyltetrazolium bromide
b a
both cell lines (P < 0.0001). Damage frequencies and tail length have statistically different at 24 h for both cell lines. Average comet density was statistically significant at 4, 8, and 24 h in MCF‑7 cells, whereas it was different at 2 and 24 h in MDA‑MB‑231 cells. The percentage of tail DNA was statistically significant at 24 h in both of them [Table 3]. The extract of S. kronenburgii may be considered as highly genotoxic against breast cancer cells. Because S. kronenburgii is an endemic plant, we could not find any data belong to this plant regarding as apoptotic and genotoxic effects.
We monitored the pyknosis which is a hallmark of apoptosis in certain cells for both cell lines after treated with S. kronenburgii extract [Figure 2] by fluorescence microscope. We suggest that treatment with S. kronenburgii extract caused apoptosis/necrosis in MCF‑7 and MDA‑MB‑231 cells for 48 h.
CONCLUSION
S. kronenburgii, endemic in Turkey, is a promising natural source for cancer therapy. The presence of distinct genotoxic Table 3: Results of comet analysis in MCF-7 and MDA-231cell lines treated with Salvia kronenburgii extract, positive and negative controls (mean±standard deviation)
Cell
type Treatment time (h) lengthComet a Average comet densitya Tail length
a Percentage
tail DNAa momentaOlive tail Percentage head DNAa GDI
a Percentage damaged cella MCF-7 0 46.04±0.99 0.21±0.00 5.22±0.25 2.21±0.16 0.46±0.03 97.79±0.16 0.15±0.01 0.00±0.00 2 48.91±1.04 0.20±0.00 6.72±0.41 2.18±0.17 0.49±0.04 97.82±0.17 0.16±0.01 0.00±0.00 4 51.20±1.15* 0.13±0.00*** 6.99±0.66 2.78±0.34 0.68±0.10 97.22±0.34 0.31±0.01*** 0.02±0.00 8 50.15±1.19 0.15±0.00*** 8.61±0.58** 4.09±0.44** 0.91±0.10* 95.91±0.44** 0.58±0.01*** 0.04±0.01*** 24 44.23±3.11 0.16±0.01*** 16.25±3.33*** 15.18±3.53*** 5.29±1.70*** 84.82±3.53*** 1.16±0.04*** 0.32±0.02*** MDA 0 39.68±0.61 0.12±0.00 4.69±0.19 2.14±0.15 0.39±0.03 97.86±0.15 0.18±0.01 0.00±0.00 2 33.25±0.63* 0.14±0.00** 3.76±0.13** 2.35±0.11 0.35±0.02 97.65±0.11 0.14±0.01 0.00±0.00 4 36.72±0.93 0.12±0.00 5.04±0.26 3.52±0.69 0.54±0.07 96.47±0.69 0.29±0.01*** 0.00±0.00 8 35.44±0.54* 0.12±0.0 4.52±0.13 3.22±0.22 0.50±0.03* 96.78±0.22 0.50±0.02*** 0.00±0.00 24 35.16±2.98 0.16±0.01*** 10.07±2.96* 9.98±2.27*** 2.98±1.42* 90.02±2.27*** 1.12±0.03*** 0.22±0.02*** EMS 24 110.30±17.22 94.31±9.25 56.82±4.54 5.92±1.33 0.13±0.03 3.69±1.25 3.88±0.04 0.97±0.02
asignificance of Salvia kronenburgii extract compared with 0 time point, *P<0.05, **P<0.005, ***P<0.0001. SE=Standard error, EMS=Ethyl methanesulfonate as positive control, GDI=Genetic damage index
Figure 2: Fluorescence imaging for determination of cell death mode. The cells were treated with 100 μg/ml Saliva kronenburgii extract for 48 h and then stained as explained in the Materials and Methods, followed by the visualization by a fluorescence microscope. Upper panel shows Hoechst33342 staining while middle and bottom panels show Calcein-AM and propidium iodide staining, respectively. Long arrows show the pyknotic/fragmented nucleus of apoptotic cells. Short arrows show the bleb formations seen in apoptotic cells (×100)
and cytotoxic activities of S. kronenburgii extract determined in this study also point out the importance of dose selection if the plant or derived compounds are intended to be used for medicinal purposes.
Acknowledgments
We thank the Research Fund of Uludağ University for the project numbered BUAP (F)‑2014/3 for providing us with the kits/chemicals. We would like to thank Buse Cevatemre for technical assistance.
Financial support and sponsorship
Nil.
Conflict of interest
There are no conflicts of interest.
REFERENCES
1. American Cancer Society. Cancer Facts & Figures 2015. Atlanta: American Cancer Society; 2015.
2. Walker JB, Sytsma KJ, Treutlein J, Wink M. Salvia (Lamiaceae) is not monophyletic: Implications for the systematics, radiation, and ecological specializations of Salvia and tribe mentheae. Am J Bot 2004;91:1115‑25.
3. Hedge IC. A Global Survey of the Biogeography of the Labiatae. In: Harley RM, Reynolds T, Eds., Science Royal Botanical Gardens, Kew; 1992. p. 7‑17.
4. Schwarz K, Ternes W. Antioxidative constituents of Rosmarinus officinalis and Salvia officinalis. II. Isolation of carnosic acid and formation of other phenolic diterpenes. Z Lebensm Unters Forsch 1992;195:99‑103.
5. Tosun M, Ercisli S, Sengul M, Ozer H, Polat T, Ozturk E, et al. Antioxidant properties and total phenolic content of eight Salvia species from Turkey. Biol Res 2009;42:175‑81.
6. Kamatou GP, Makunga NP, Ramogola WP, Viljoen AM. South African Salvia species: A review of biological activities and phytochemistry. J Ethnopharmacol 2008;119:664‑72.
7. Topçu G, Türkmen Z, Schilling JK, Kingston DG, Pezzuto JM, Ulubelen A. Cytotoxic activity studies on some Anatolian Salvia extracts. Pharm Biol 2008;46:180‑4.
8. Tayarani‑Najaran Z, Asili J, Aioubi E, Emami SA. Growth inhibition and apoptosis induction of Salvia chloroleuca on MCF‑7 breast cancer cell line. Iran J Pharm Res 2013;12:789‑99.
9. Li C, Han X, Zhang H, Wu J, Li B. The interplay between autophagy and apoptosis induced by tanshinone IIA in prostate cancer cells. Tumour Biol 2016;37:7667‑74.
10. Zhang K, Li J, Meng W, Xing H, Yang Y. Tanshinone IIA inhibits acute promyelocytic leukemia cell proliferation and induces their apoptosis in vivo. Blood Cells Mol Dis 2016;56:46‑52.
11. Lang JY, Chen H, Zhou J, Zhang YX, Zhang XW, Li MH, et al. Antimetastatic effect of salvicine on human breast cancer
MDA‑MB‑435 orthotopic xenograft is closely related to rho‑dependent pathway. Clin Cancer Res 2005;11:3455‑64.
12. Jantová S, Hudec R, Sekretár S, Kučerák J, Melušová M. Salvia officinalis L. extract and its new food antioxidant formulations induce apoptosis through mitochondrial/caspase pathway in leukemia L1210 cells. Interdiscip Toxicol 2014;7:146‑53.
13. Russo A, Formisano C, Rigano D, Senatore F, Delfine S, Cardile V, et al. Chemical composition and anticancer activity of essential oils of mediterranean sage (Salvia officinalis L.) grown in different environmental conditions. Food Chem Toxicol 2013;55:42‑7. 14. Zhao Q, Huo XC, Sun FD, Dong RQ. Polyphenol‑rich extract of Salvia
chinensis exhibits anticancer activity in different cancer cell lines, and induces cell cycle arrest at the G0/G1‑phase, apoptosis and loss of mitochondrial membrane potential in pancreatic cancer cells. Mol Med Rep 2015;12:4843‑50.
15. Davis PH, Tan K, Mill RR. Flora of Turkey and the East Aegean lslands. Vol. 10. Edinburgh: Edinburgh University Press; 1988.
16. Ulukaya E, Ozdikicioglu F, Oral AY, Demirci M. The MTT assay yields a relatively lower result of growth inhibition than the ATP assay depending on the chemotherapeutic drugs tested. Toxicol In Vitro 2008;22:232‑9.
17. Andreotti PE, Cree IA, Kurbacher CM, Hartmann DM, Linder D, Harel G, et al. Chemosensitivity testing of human tumors using a microplate adenosine triphosphate luminescence assay: Clinical correlation for cisplatin resistance of ovarian carcinoma. Cancer Res 1995;55:5276‑82.
18. Singh NP, McCoy MT, Tice RR, Schneider EL. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp Cell Res 1988;175:184‑91.
19. Anderson D, Yu TW, Phillips BJ, Schmezer P. The effect of various antioxidants and other modifying agents on oxygen‑radical‑generated DNA damage in human lymphocytes in the COMET assay. Mutat Res 1994;307:261‑71.
20. Oztekin N, Başkan S, Evrim Kepekçi S, Erim FB, Topçu G. Isolation and analysis of bioactive diterpenoids in Salvia species (Salvia chionantha and Salvia kronenburgiii) by micellar electrokinetic capillary chromatography. J Pharm Biomed Anal 2010;51:439‑42. 21. Lim H, Son KH, Chang HW, Kang SS, Kim HP. Effects of anti‑inflammatory
biflavonoid, ginkgetin, on chronic skin inflammation. Biol Pharm Bull 2006;29:1046‑9.
22. Abu‑Dahab R, Abdallah MR, Kasabri V, Mhaidat NM, Afifi FU. Mechanistic studies of antiproliferative effects of Salvia triloba and Salvia dominica (Lamiaceae) on breast cancer cell lines (MCF7 and T47D). Z Naturforsch C 2014;69:443‑51.
23. Zare Shahneh F, Valiyari S, Baradaran B, Abdolalizadeh J, Bandehagh A, Azadmehr A, et al. Inhibitory and cytotoxic activities of Salvia officinalis L. extract on human lymphoma and leukemia cells by induction of apoptosis. Adv Pharm Bull 2013;3:51‑5.
24. Godard T, Deslandes E, Lebailly P, Vigreux C, Sichel F, Poul JM, et al. Early detection of staurosporine‑induced apoptosis by comet and annexin V assays. Histochem Cell Biol 1999;112:155‑61.
25. Dhivya R, Jaividhya P, Riyasdeen A, Palaniandavar M, Mathan G, Akbarsha MA, et al. In vitro antiproliferative and apoptosis‑inducing properties of a mononuclear copper(II) complex with dppz ligand, in two genotypically different breast cancer cell lines. Biometals 2015;28:929‑43.