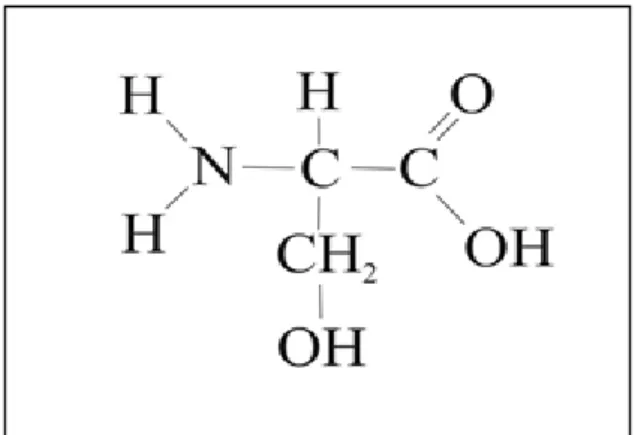
ÇINAR AĞAÇLARINDA (Platanus orientalis L.) YAŞA BAĞLI OLARAK BAZI KİMYASAL BİLEŞENLERİN BELİRLENMESİ
Tam metin
Şekil
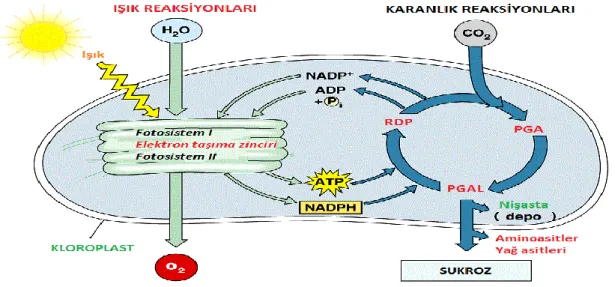


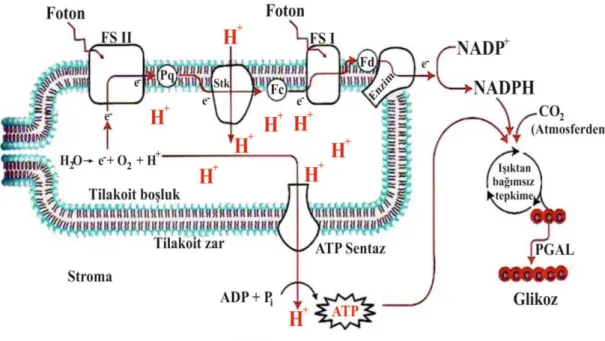
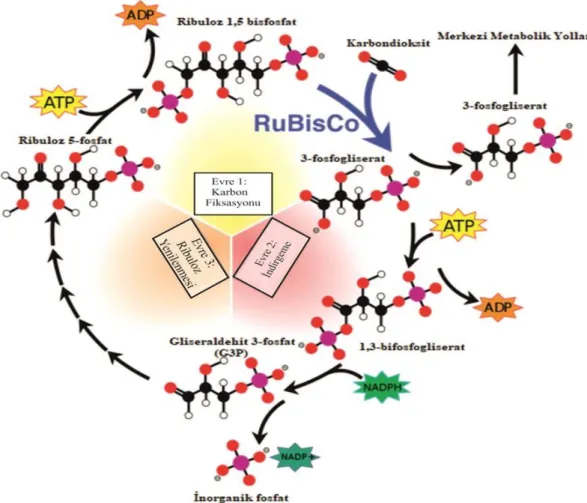

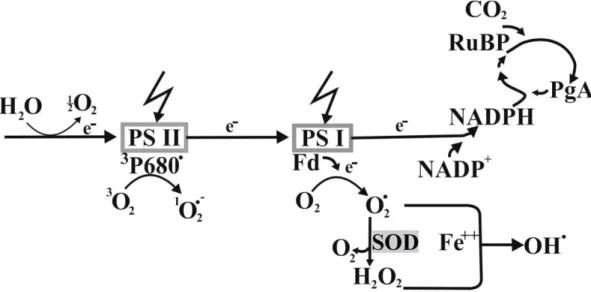
Outline
Benzer Belgeler
Konser alanında toplam 1960 kişilik olduğuna göre, en arka sırada kaç
Kırk sekiz aylık yaş dönemindeki İngiliz atlarında bacak konformasyon parametrelerine cinsiyetin etkisi değerlendirildiğinde; dişilerde metacarpus açısı (P<0,001)
Lider davranışları, liderin etkileşim alanındaki kişilerin motivasyonlarında, kültür oluşturmada ve yapıyı şekillendirmede önemli rol oynar.Örgütlerde insan
[r]
[r]
Fonksiyonlar: Fonksiyonlar, birebir, üzerine, eşit, birim fonksiyon, morfizm ve konu ile ilgili soruların çözülmesi; Sayılar: Reel sayılar, Tamsayılarda bölünebilme ve konu
Mezun
MÜHENDİSLİK FAKÜLTESİ ELEKTRİK ELEKTRONİK MÜHENDİSLİĞİ PROGRAMI 2021 - 2022 ÖĞRETİM YILI ÖĞRETİM PLANI (MÜFREDAT). İSTANBUL COMMERCE UNIVERSITY FACULTY OF ENGINEERING
