Sorgum (Sorghum bicolor (L.) Moench) koleksiyonunda sorgum afitine (Melanaphis sacchari (Zehntner) dayanıklılık ve uzun boy karakterlerinin moleküler markerler yardımıyla belirlenmesi
Tam metin
Şekil




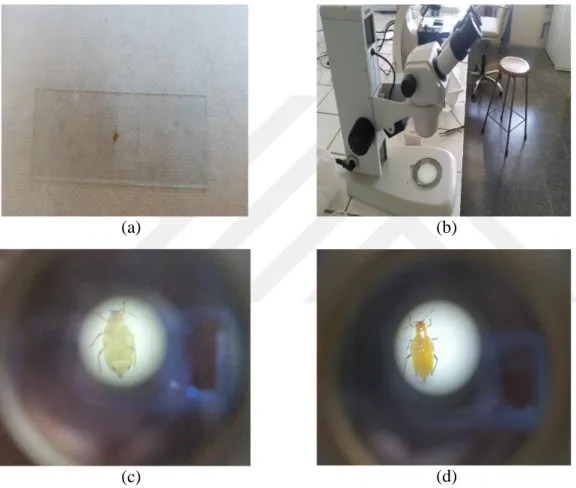


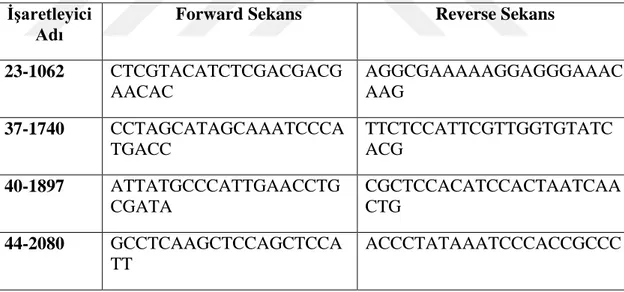
Benzer Belgeler
Olgumuzda hipertioidi ve TRAB pozitifliği olması Graves açısından değerlendirilebile- cek bulgulardır, bununla birlikte olgumuzda ekzoftal- mus ve pretibial miksödem gibi
In univariate analysis, we found that patients with brain metastases from lung cancer with age ≤60 years, small-cell ca, the absence of brain metastasectomy, controlled
• Aktivitenin özelliğine göre kasların kasılma şekline göre dinamik ve statik dayanıklılık.. • Aktivitenin süresi bakımından kısa, orta ve uzun süreli dayanıklık
While the military inse- curities that Turkey has experienced and the specific foreign policy acts designed in response to them have been accounted for in the literature,
Kendi bedeninin yerini tanımlamak, kendine bir yer bulmak, şair olarak sayfada kendine bulduğundan farklı bir yer bulmak için Acconci daha önceden ilgilendiği şiirden
Yapılan istatistiksel analiz sonucunda vücut ağırlığı, vücut sıvı miktarı, kas kütlesi, vücut yağ yüzdesi ve vücut sıcaklığı değerlerinde anlamlı fark
Beydar ı sorgum çe şidinin üç ekim zaman ı ve üç bitki sı kl ığı nda oluşturdukları çeş itli agronomik özelliklere ili şkin varyans analizi sonuçlar ı Çizelge 4 ve
Araştırmada kullanılan sorgum sudan otu melezi çeşitlerinin farklı leonardit uygulaması dozlarındaki ADL içeriğine ilişkin varyans analiz sonuçları Çizelge
