©BEYKENT UNIVERSITY
EFFECTS of PARASITISM by
ENDOPARASITOID Pimpla turionellae on
HEMOLYMPH PROTEINS of HOST
Galleria mellonella
Süleyman KALELI
1*, M. Yaşar AKSOYLAR
2,
Abdurrahman AKTUMSEK
3, C. COKMUS
41
Department of Biology, Isparta Healty College, Suleyman Demirel
University, Isparta, Turkey
2
Department of Biology, Sciences and Arts Faculty, Suleyman
Demirel University, Isparta, Turkey
3
Department of Biology, Sciences and Arts Faculty, Selcuk
University, Konya, Turkey
4
Department of Biology, Sciences Faculty, Ankara University, Ankara,
Turkey
ABSTRACT
The plasma proteins were determined under the effect of parasitism by endoparasitoid, Pimpla turionellae (Hymenoptera: Ichneumonidae) on hemolymph proteins of host, Galleria mellonella (Lepidoptera: Pyralidae) by the colorimetric and SDS-PAGE (Sodium dodecyl sulfate- polyacrylamide gel electrophoresis) methods. In the colorimetric analysis it was showed a decrease in the total plasma protein levels in parasitized host during the first hour compared with unparasitized controls whereas increased total plasma protein levels were determined during the third hour. However, In the SDS-PAGE analysis it revealed that the levels of 15.5, 17.5, 67 and 82 kDa proteins decreased in parasitized host, compared to unparasitized controls. After 72 h of treatment, plasma protein in the parasitized host was different to that of unparasitized controls. In conclusion, because of parasitization protein levels were found as a different in the colorimetric and the electrophoresis analysis. Keywords: Hemolymph protein, SDS-PAGE, parasitization, Pimpla turionellae, Galleria mellonella
Özet
Bu çalışmada Endoparasitoid Pimpla turionellae (Hymenoptera: Ichneumonidae) tarafından parazitlenmiş konak Galleria mellonella
(Lepidoptera: Pyralidae)'da parazitlemenin etkileri kolorimetrik ve SDS-PAGE (Sodium dodecyl sulfate- polyacrylamide gel electrophoresis) yöntemleri ile hemolenf proteinler araştırılmıştır. Kolorimetrik analizlerde, parazitlenmemiş kontrolle ilk saat karşılaştırıldığında parazitlenmiş konağın total plazma proteinlerinde bir azalma ve üçüncü saat süresince belirlenmiş total protein düzeylerinde ise artış görülmüştür. Bununla birlikte, SDS-PAGE analizinde, kotrollerle karşılaştırıldığında parazitlenmiş konakta 15.5, 17.5, 67 ve 82 kD protein düzeylerinin azaldığı ortaya çıkmıştır. Parazitlemeden sonraki 72. saatte parazitlenmiş konağın plazma proteini kontrole göre farklı olduğu görülmektedir. Sonuç olarak parazitlemeden dolayı protein düzeyleri kolorimetrik ve elektroforetik analizlerde farklı bulunmuştur.
Anahtar kelimeler: Hemolenf protein, SDS-PAGE, parazitleme, Pimpla turionellae, Galleria mellonella
1. INTRODUCTION
Use of parasitoids to regulate the density of a particular pest insect is an attractive way in biological control programmes and integrated pest management systems, thus creating a need to expand our knowledge of host-parasitoid relationship (DeBach and Rosen, 1991; Dorn, 1993; Ockroy et al., 1997).
Eggs of endoparasitoids are laid directly into the hosts hemocoel. Ichneumonid and braconid parasitoids co-inject eggs, venom and polydnaviruses into the host (Lavine and Beckage, 1995; Strand and Pech, 1995; Shelby and Webb, 1999). During parasitization, the presence of parasitoid- derived factors evokes a striking re-programming of host behaviour, reproductive potential, development and immunity (Beckage, 1993; Lawrence and Lanzrein, 1993).
When endoparasitoid insects parasite their hosts, especially, lepidoptereous species, qualitative and quantitative changes in the host plasma protein profile often occur. In some cases, the levels and timing of appearance of naturally-occurring host proteins may be altered. For example, parasitism of
Trichoplusia ni by Hyposoter exiguae decreased the concentration of several
host hemolymph proteins (Richards and Edwards, 1999). Similarly, parasitism of Pieris rapae by Apanteles glomeratus ( Smilowitz and Smith, 1977) resulted in decrease in the concentration of host storage protein, while parasitism of Manduca sexta by A. congregatus (Beckage and Templeton,
1986) decreased the arylphorin concentration. By contrast, arylphorin levels increased in T. ni parasitized by Chelonus sp. (Kunkel et al., 1990).
Many endoparasitoids have been shown to interfere with the development of their host insect- and parasitoid- associated factors such as polydnaviruses, venom and teratocytes appear to play important roles in this respect (Lawrence and Lanzrein, 1993). Polydnaviruses have been described in certain members of the families Braconidae and Ichneumonidae of
Hymenoptera (Stoltz et al., 1995), whereas, teratocytes were found only in certain members of the families Braconidae, Platygasteridae and Sclonidae of Hymenoptera (Dahlman and Vinson, 1993).
Parasitism-specific proteins (PSPs) occur in insect hosts after attack by a variety of parasitic hymenopterous species, most notably the braconid and ichneumonid parasitoids of lepidopteran hosts (Soldevila and Jones, 1991; Harwood and Beckage, 1994; Li and Webb, 1994). A limited number of such species have been evaluated with respect to their molecular compositions or their origins (Harwood and Beckage, 1994; Li and Webb, 1994). So far PSPs from only one dipteran host species have been reported (Lawrence, 1990; Rolle and Lawrence, 1994a,b). However, their origins, molecular compositions or their functions are unknown.
In addition, new proteins which are not of host origin may sometimes appear with their site of synthesis and timing of appearance may vary or depend on the particular endoparasitoid hymenopterous species and host concerned (Beckage et al., 1987; Jones, 1989; Beckage, 1993; Beckage and Kanost, 1993).
The effect of parasitism by parasitoid species on the plasma protein profile of their insect hosts are less well known. The current study was undertaken to investigate how parasitism of host, Galleria mellonella parasitized by endoparasitoid, Pimpla turionellae.
2. MATERIALS AND METHODS
2.1. Insects
Hosts, greater wax moths, Galleria mellonella L. (Lepidoptera: Pyralidae) were mass cultured at 25 oC, 60 % relative humidity and 12 hours light: 12 hours dark on semiartificial diet (Bronskill, 1961). Pimpla turionellae L. (Hymenoptera: Ichneumonidae) is a solitary endoparasitoid which can develop successfully in Galleria pupae. Therefore, they were reared routinely on pupae of G. mellonella in insect laboratory of Biology Department, Sciences and Arts Faculty, Selcuk University, Konya, Turkey. Control and parasitized insects were held under the same conditions.
2.2. Hemolymph collection
G. mellonella pupae were divided into two groups, namely a)
parasitized pupae: G. mellonella pupae were parasitized by mature females of endoparasitoid P. turionellae, b) unparasitized pupae (control). Hemolymph samples were taken at 1, 3, 6, 12, 24, 36, 48, 60 and 72 after parasitism. Samples were collected by cutting at abdomen and drawing into glass capillary tubes. Samples were pelletted at 5000 g for 4 minutes at 4 oC and stored at -18 ° C until use. A small amount of phenylthiourea was added into
all eppendorf tubes for hemolymphs collected to prevent melanisation and oxidation (Zupko et al., 1993).
2.3. Electrophoresis
Hemolymph samples were removed from freezer, diluted in a buffer containing 0.05 M Tris-HCl (pH 6.8) and centrifuged at 12.500 g for 10 minutes. Supernatant and SDS sample buffer were mixed in the ratio of 3:1 followed by denaturation of the proteins in boiling water for 5 minutes.
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) was performed as described by Laemmli (1970), using 4% (w/v)
stacking and 10 % (w/v) resolving gel. Protein bands were visualized by
Coomassie Brillant Blue R250 (Sigma). The molecular weight of each protein band could then be calculated according to the standard curve of purified marker proteins: • lactalbumin cow's milk (14.2 kDa), trypsin inhibitor -soybean (20.1 kDa), carbonic anhydrase - bovine erythrocytes (29 kDa), ovalabumin egg (45 kDa), albumin bovine (66 kDa), phosphorylase B -rabbit muscle (97.4 kDa), • -galactosidase - E. coli (116 kDa), myosin - -rabbit
muscle (205 kDa) from Sigma (Cat. No. M. 2789). The current study was
carried out using Technicon RA-XT autoanalyzator. 2.5 ^l protein extract was loaded into each well of gels and the electrophoresis was run from cathode to anode for 1 h at 20 mA and 3 h at 35 mA at 4 °C.
The semi-quantification of the protein concentration of the samples was determined by using the method of Esen (1978). The protein extracts were applied as 2.5 ^l onto the surface of a set Whatmann paper (No 1) cut out as bands and were dried.
2.4. Statistical Analysis
Experiments were repeated at least three times. All data were analysed using a computerized statistical package (MSTATC).
3. RESULTS
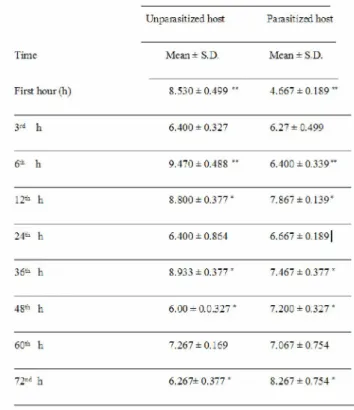
Total plasma protein values of host, Galleria mellonella pupae were clearly altered by the parasitism of endoparasitoid Pimpla turionellae (Table 1).
There was a dramatic decrease in the total protein concentration on the first hour of the parasitized host following the parasitism (p < 0.01). Similarly, a dramatic decrease was also observed on the 6th hour after parasitism. However, decreases during the 12th and 36th hour were significant (p < 0.05). On the other hand, the total plasma protein concentrations of parasitized host were increased during the 48th and 72th hour (p < 0.05), although, no significant changes were observed during the 3rd, 24th and 60th hour between total plasma protein levels of parasitized and unparasitized hosts (p > 0.05).
The proteins with size of > 205, 46, 42, 36, 34, 26 and 18 kDa decreased in parasitized host compared to controls (Fig. 1). Proteins at 36 and 34 kDa sizes decreased remarkably after the 24 hours, and 36 kDa sizes disappeared from the host after 72 hours parasitism. In contrast, protein at 36 kDa size remarkably increased on unparasitized host on 72 hours following parasitism. Proteins at 67 and 82 kDa sizes were easily visualised on the gel as a single band due to their higher densities. Protein at 82 kDa size decreased until the 24th hours and increased after the 24th hours in parasitized host. Protein at 67 kDa size was found to decrease in the first hour but it was seen to increase after the first hour in parasitized host. Proteins at 17.5 and 15.5 kDa sizes decreased until the 24th hour and increased in parasitized host thereafter.
Table 1. Total plasma protein values at different time of parasitized by endoparasitoid P.
turionellae and unparasitized host G. mellonella pupae.
Total plasma proteins of "host fp'dl)1
Unparasitized host Parasitized "host
Time Mean± S.D. Mean = S.D.
First hour (h) 8.530 - 0.466 '" 4.667 = 0.186 "" 3'J h 6.400 - 0.327 6.27 = 0.466 6^ i 6.470 - 0.488 " 6.400 = 0.336™ 12a h 8.800 = 0.377 " 7.867 = 0.136" h 6.400 = 0.864 6.667 = 0.186| 36^ h 8.933 = 0.377 " 7.467 = 0.377" 48a h 6.00 = 0.0.327 " 7.200 = 0,327 " 60th h 7.207 = 0.166 7.067 = 0.754 72"d h 6.267= 0.377 " 8.267 = 0.754 "
!Hie average of three Experiments. *: p<0.05, ** : p<0.01
Figure 1. Protein profiles of pupae hemolymph proteins of G. Mellonella parasitized by P.
turionellae and unparasitized host G. mellonella at different periods. (Lane 2, 4, 6, 8, 10, 12, 14, 16 and 18 are parasitized hosts; lane 3, 5, 7, 9, 11, 13, 15, 17 and 19 are unparasitized hosts; Lane 1 and 20 are the molecular weight standards. And lanes 2-3 at first h, lanes 4-5 at 3rd h,
lanes 6-7 at 6h h, lanes 8-9 at 12th h, lanes 10-11 at 24th h, lanes 12-13 at 36th h, lanes 14-15 at
48th h, lanes 16-17 at 60h h, lanes 18-19 at 72nd h).
4. DISCUSSION
Earlier studies showed that the total amount of hemolymph protein is reduced during parasitism, beginning almost immediately after the host is parasitized. Simultaneously, parasitism induces synthesis of large amounts of novel proteins that appear in the blood as early as 1-2 hours post-parasitization. The present report confirms earlier studies describing the presence of novel proteins in larvae and pupae, and also characterizes the effects of parasitism in altering the titers of several endogenous host hemolymph proteins normally produced by the fat body and other tissues (Beckage and Kanost, 1993). Previous reports presented the physiological arrestment and alterations in hormone and hemolymph protein titers of lepidopteran larvae of permissive host following parasitization by Euplectus species (Coudron et al., 1990, 1994).
Successful endoparasitic development requires suppression of the humoral system, but not its irreversible shutdown. The invading parasitoid has to ensure that it is not destroyed by the immune processes, but that immuno-compromised host maintains the ability to fight secondary infection. It is critical that the immune reaction is not undermined to such an extent, that death of both host and developing parasitoid cannot be avoided (Ockroy et al., 2002).
Invasion by parasitoids or by bacteria induces arrays of new hemolymph proteins in the host insects (Beckage et al., 1989; Soldevila and Jones, 1993; Rolle and Lawrence, 1994a). In Pieris brassicae upon parasitism by Cotesia glomerata also considerable changes in the hemolymph protein pattern occurs. These changes, however, were clearly distinct from hemolymph protein patterns obtained after bacterial infection on wounding. For example, a new 38 kDa protein was found in P. brassicae two days after oviposition of parasitic wasp (Ockroy et al., 1997). New proteins appears in the hemolymph at defined times post-oviposition. It is suggested that such proteins have a function linked to alterations in behaviour, development and immunity of the host (Ockroy, 1999).
Many studies demonstrating that parasitization by Cotesia congregata induces synthesis of novel hemolymph proteins that begins immediately after the host is parasitized, specifically to show that titers of endogenous host proteins also are dramatically affected by parasitism (Beckage et al., 1987, 1989; Harwood, 1993).
Parasitism-specific proteins have been detected in numerous endoparasitoid-host model systems, their sources are variable, and many help to condition the host to ease parasitism. For example "new" host proteins might be the result of parasitoid-mediated rediction of host protein synthesis, which may make the host plasma more nutritious for the endo- or ectoparasitoid (Thompson, 1993; Coudron et al., 1994). Alternatively, many parasitism-specific proteins are the products of polydnavirus genes, which suppress and distrupt the host immune and endocrine systems, respectively (Cook et al, 1984; Vinson, 1990a; Beckage, 1993, 1997, 1998; Strand and Pech, 1995; Harwood et al., 1998).
In the study confirms that protein profiles of hemolymph of host insects affected by parasitism. Results from this experiments indicated that 72 hours of treatment, plasma from parasitized pupae was different to that unparasitized controls. At the end of first hour of parasitization, table and figure showed that total protein levels and protein profiles in parasitized hosts were dramatically decreased. All proteins were reduced by parasitoid at the beginning of the first hour after parasitization. > 205, 46 and 42 kDa proteins, and 36, 34, 26 and 18 kDa proteins at the following hours were reduced. Titers of various hemolymph proteins in parasitized hosts were increased in the different times. For example, 67 kDa protein after 3 hours, 15.5 and 17.5 kDa proteins after 24 hours and 82 kDa protein after 36 hours were different than controls. 36 and 34 kDa are probably major protein or larval specific protein because of these molecular weight protein was exist in both of host and endoparasitoid hemolymph protein profiles. Beckage et al (1989) reported that 33 kDa proteins on host-parasitoid systems were, also, seen after 3 hours of parasitizing the host, Manduca sexta by parasitoid C. congregata. However,
it was reported that the reason of the formation of 33 kDa proteins in parasitized M. sexta hemolymph was the infection of polydnavirus which was secreted from ovarian calyx of C. congregata. Luckhart and Webb (1996) pointed that polydnavirus did not occur in ovarian calyx, but 29-36 kDa proteins groups in ovarian calyx fluid were found. These proteins were present from 3 to 96 hours in parasitism of parasitized host hemolymph, and are reported to be in the form of the glycoform. In this study, new proteins were not seen due to parasitization, suggesting that there was no virus secreted ovarian calyx of P. turionellae. This situation, also, supports the study of Luckhart and Webb (1996). According to the study of parasitized hosts were shown in A. suspense pupae at the 24th and 36th, and this protein was described to be a specific protein bound to parasitism. Roll and Lawrence (1994b) reported the 24 kDa protein was in the nature of a glycoprotein. Glycoproteins left into host by parasites can induce the formation of patogenity since they can potentially be dangerous factor to host and glycoproteins are the sources of nutrient for parasite feedings.
In conclusion, the hemolymph proteins were affected in parasitized host as seen in the table and the figure. The study shows the effects of relation between parasite and host. We think that the effects of parasite on host may be studied as amino acid and hormone levels in the future.
REFERENCES
Beckage, N.E., Templeton, T.J.; Physiologial effect pf parasitism by
Apanateles congregatus in terminal stage tobacco hornworm larvae. Journal
of Insect Physiology, 32 (1986), 299.
Beckage, N. E., Nesbit, D. J., Nielsen, B. D., Spence, K. D., Barman M. A.E.; Alteration of hemolymph polypeptides in larvae parasitized by Cotesia
congregata: a two-dimensional electrophoretic analysis and comparison with
major bacteria induced proteins. Archs Insect Biochem. Physiol. 10 (1989), 29-45.
Beckage N, E., Templeton T, J., Nielsen B, D., Cook D, I., Soltz D, B.; Parasitism-induced haemolymph proteins in Manduca sexta (L.) larve parasitized by the braconid wasp Cotesia congregata (Say). Insect. Biochem.,
17 (1987), 439-455.
Beckage, N.E.; Games parasites play: The dynamic roles of proteins and peptides in the relationship between parasite and host. In: Beckage. N.E., Thompson, S.N., Frederci, B.A. (Eds.). Parasites And Pathogens of Insects, vol. 1. Academic Press, New York, (1993), 25-57.
Beckage, N.E.; New insights: How parasites and patogens alter the endocrine physiology and development of insects hosts. In:Beckage, N.E.(Ed.). Parasites And Pathogens. Effect On Host Hormones And Behaviour. Chapman and Hall, London (1997), 3-36.
Beckage, N.E.; Modulation of immune respones to parasitoids by polydnaviruses. Parasitology, 116 (1998), S57-S64.
Beckage, N.E., Kanost, M.R.; Effect of parasitism by the braconid wasp
Cotesia congregata on host hemolymph proteins of the tobacco hornworm Manduca sexta. Insect Biochemistry and Molecular Biology 23 (1993),
643-653.
Bronskill, J.K.; A cage to simplify the rearing of the greater wax moth
Galleria mellonela (Pyralidae). J. Lep. Soc. (1961) 102-104.
Cook, D.I., Stoltz, D.B., Vinson, S.B.; Induction of a new haemolymph glycoprotein in larvae of a permissive hosts parsitized by Campoletis
sonorensis. Insect Biochemistry 14 (1984), 45-50.
Coudron, T.A., Jones, D., Jones, G.; Premature production of late larval storage proteins in larvae of Trichoplusia ni parasitized by Euplecturus
comstockii. Arch Insect Biochem Physiol, 26 (1994), 97-109.
Dahlman, D. I., Greene, J. R.; Larval hemolymph proteins in tobacco hornworms parasitized by Apanteles congregatus. Ann. Ent. Soc. Am., 74 (1981), 130-133.
Dahlman, D.I., Vinson, S.B.; Teracytes: developmental and biochemical characteristics. In: Beckage, N.E., Thomson, S.N., Federici, B.A. (Eds). Parasites and patogens of Insect, vol.1. Academic Press, San Diego (1993) 145-165.
DeBach, P., Rosen, D.; Biological Control by Naturel Enemies. Cambridge University Press, Cambridge, UK (1991).
Dorn, S.; Systembezogene Schadlingsbekampfung. Landwirtschaft Shweiz 6 (1993), 385-389.
Esen, A.; A simple method for quantitative, semiquantitative and qualitative assay of protein. Anal. Biochem. 89 (1978), 264-273.
Harwood S. H.; A Cotesia congregata polydnavirus gene cloning, characteriztation, and pattern of expression in host sphingid larvae. Ph.D. Dissertation, University of California-Riverside (1993).
Harwood, S., Beckage, N.; purification characterization of an abundant polydnavirus-induced protein from the hemolymph of the Manduca sexta larvae parasited by Cotesia congregata. Insect Biochem. Biol. 24 (1994),685-698.
Harwood, S.H., McElfresh, J.S., Nguyen, A., Conlan, C.A., Beckage, N.E.; Production of early expressed parasitisim-specific proteins in alternate sphingid hosts of the braconid wasp Cotesia congregata. Journal of Invertebrate Pathology 71 (1998), 271-279.
Jones, D.; Protein expression during parasite redirection of host (Trichoplusia
Kunkel, J.G., Grossniklaus-Burgin, C., Karpells, S.T., Lanzrein, B.; Arylphorin of Trichoplusia ni: characterization and parasiteinduced precocious increase in titre. Archives of Insect Biochemistry and Physiology 13 (1990), 117-125.
Laemmli, U.K.; Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227 (1970), 680-685.
Lavine, M.D., Beckage, N.E.; Polydnaviruses: potent mediators of host insect immune dysfunction. Parasitol. Today 11 (1995), 368-378.
Lawrence, P.O.; Release of polypeptides from serosal cell of Biosteres
longicaudatus (Hym: Braconidae). Arch. Insect Biochem. Physiol. 13 (1990),
199-216.
Lawrance, P.O., Lanzrein, B.; Hormonal interactions between insect endoparsites and their host insects. In:Beckage, N.E., Thomson, s.n., Federici, B.A.,(Eds). Parasites and Pathogens of Insect. Vol.1. Academic Press. San Diego. (1993), 59-86.
Li, X., Webb, B.; Apparent functional role for cysteine-rich polydnavirus protein in suppression of the insect cellular immune response. J. Virol. 68 (1994), 7482-7489.
Luckhart, S., Webb, B.A.; Interaction of a wasp ovarian protein and Polydnavirus in host immune suppression. Develop. And Comp. Immunol. 20 (1996), 1-21.
Ockory, K.; Eine Parasitierung durch die Schlupfwespe Cotesia glomerata L. (Hymenoptera:Braconide) führt, in Abhangigkeit zur endoparasitischen Entwicklung der Wespe, zu Veranderungen im Hamolymph-Proteinspektrum des Wirtes Pieris brassicae L. (Lepidoptera: Pieridae). Ph.D. Thesis, ETH Zurich, Switzerland (1999).
Ockroy, K., Trenczek, T.E., Dorn, S.; Parasitism by Cotesia glomerata induces several changes in the hemolymph protein pattern of the host insect
Pieris brassicae. Verh. Dtsch. Zool.Ges. 90 (1997),130.
Ockroy, K.S., Cole, T.J., Trenczek, T.E., Doron, S.; Comparsion of parasitism by Cotesia glomerata with bacterial infection and wouding in Pieris brassicae: induction of new haemolymph polypeptides and changes in humoral immune response. Journal of Invertebrate Patology 81 (2002), 12-18.
Richards, E.H., Edwards, J.P.; Parasitization of Lacanobia oleracea (Lepidoptera) by the ectoparasitic wasp, Eulophus pennicornis, supp (1999). Rolle, R.S., Lawrence, P.O.; Characterization of a 24 kDa parasitism-specific hemolymph protein from pharate pupae of the Caribbean furit fly Anastrepha
suspense .Arch. Insect Biochem. Physiol. 25 (1994a), 265-285.
Rolle, R.S., Lawrence, P.O.; Purification of a 24 kDa parasitism-specific hemolymph protein from pharate pupae of the Caribbean furit fly Anstrepha
suspense. Arch. Insect Biochem. Physiol. 27 (1994b), 265-285.
Shelby, K,S., Webb, B.A.; Polydnavirus - mediated suppression of insect immunity. J.Insect Physiol. 45 (1999), 507-514.
Smilowitz, A., Smith, C.L.; Hemolymph proteins of developing Pieris rapae larvae parasitized by Apanteles glomeratus. Annual Entomologial Society of America 70 (1977), 447-454.
Sodevila, A.J., Jones, D.; Expression of a parasitism-specific protein in Lepidopteran hosts of Chelonus sp. Arch. Insect Biochem. Physiol. 24 (1993), 149-169.
Soldevila A. I., Jones D.; Immunoanalysis of unique protein in Trichoplusia ni larvae parasitized by the braconid wasp Chelonus near Curvimaculatus. Insect Biochem. 21(1991), 845-856.
Stoltz, D.B., Beckage, N.E., Bilissard, G.W., Fleming, J.G.W., Krell, P.J., Theilmann, D.A., Summers, M.D., Webb, B.A.; Polydnaviridae. In. Murphy, F.A., Fauqent, C.M., Bishop D.H. (Eds), Virus Taxonomy: 6th Report of the International Committee of Viruses. Springer Verlag, Wien and New York (1995), 143-147.
Strand, M.R., Pech, L.L.; Microplitis demolitor polydnavirus induces apoptosis of aspecific hemocyte morphotype in Pseudoplusia includens. J. Gen. Virol. 76 (1995), 283-291.
Thompson, S.N.; Redirection of host metbolism and effect on parasite nutrition. In:Beckage, N.E.,Thompson, S.N., Federici, B.A. (Eds.). Parasites and Pathogens of Insect, vol. I, Academic Press, London, (1993), 125-144. Vinson, S.B.; Physiological interactions between the host genus Heliothis and its guild of parasitoids.arch Insect Biochem Physiol. 13 (1990a), 63-81.
Zupko, K., Sklan, D., Lensky, Y.; Proteins of the honeybee (Apis mellifera L.) body surface and exocrine gland secretions. J. Insect Physiol. 39 (1993), 41-46.