Türkiye'de yetiştirilen bazı bodur fasulye (Phaseolus vulgaris L.) genotiplerinin bor ve çinko uygulamalarına tepkilerinin belirlenmesi
Tam metin
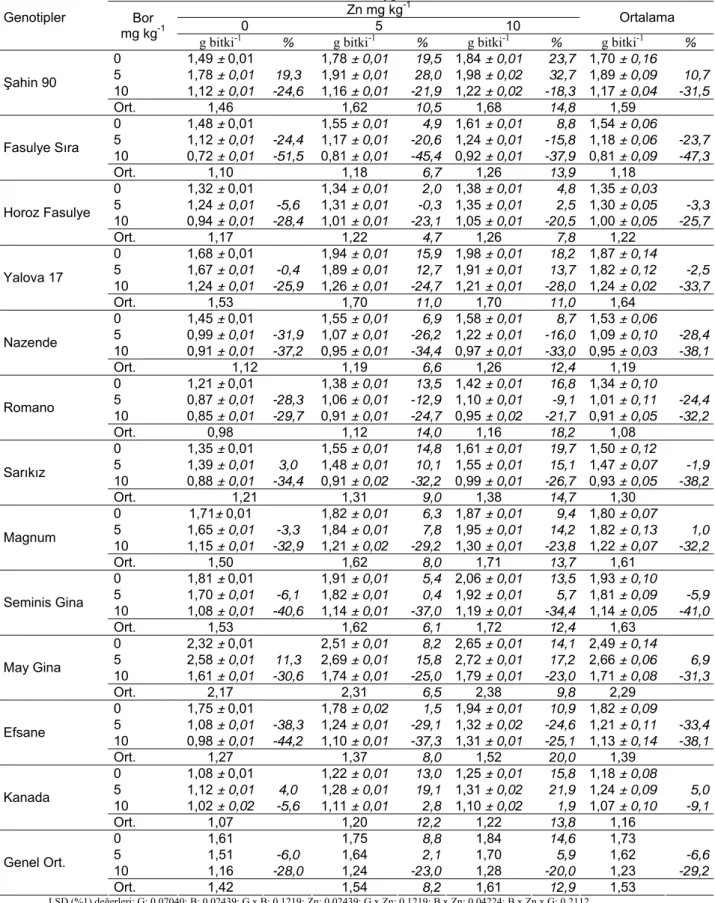
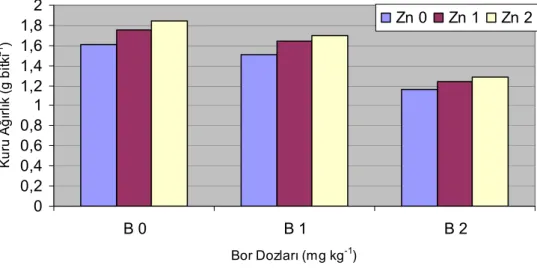
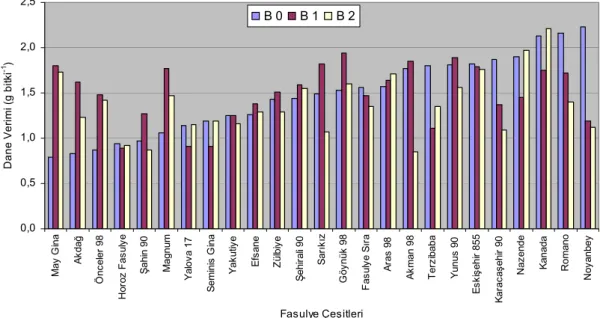
Şekil


Outline
Benzer Belgeler
Piyanist Gülsin Onay, önümüz deki yıl Danim arka'nın başkenti Kopenhag, Alborg ve Arhus gibi kentlerinde konserler vermek üzere. davet
[r]
The main objective of this study was to estimate the shielding parameters of glasses consisted of CdO–WO 3 –TeO 2 ternary system by using WinXCom software
As the second case of the six-sigma methodology, we consider its implementation in the process management and logistic. The methodology has been conducted in an international
Çalışma sonucunda, presleme tekniğinin tabakalama tekniğine göre daha yüksek bir bağlanma dayanımı sağladığı, liner materyalinin hem tabakalama hem de presleme
276 “Kar Zarar Ortaklığı Belgelerinin Kurul Kaydına Alınmasına İlişkin Esaslar Tebliği” Seri: III, No: 27 R.. Piyasası Kurulu’nca belirlenen rakamla çarpılır,
Son yıllar yeni tüketim formlarının hayatımıza girmesine sahne olmuş, ortaya çıkan tüketim toplumu/kültürü kavramsallaştırmaları beraberinde yeni ilişkiler
Geçmişi çok karanlık, Parisli bir hayat kadını olan Anjel, ahlâklı, dindar ve namuslu Matmazel Anjel olarak, Dehri Efendi’nin konağına mürebbiyelik etmek için girer..
