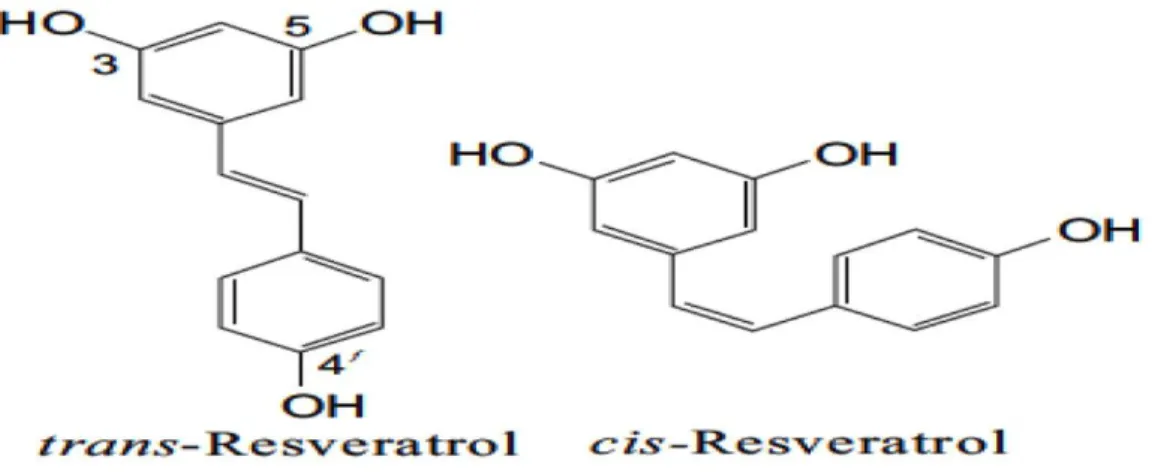
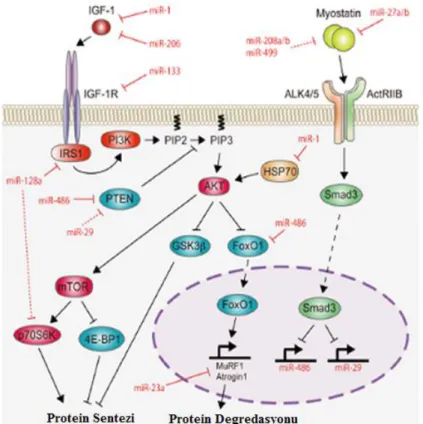
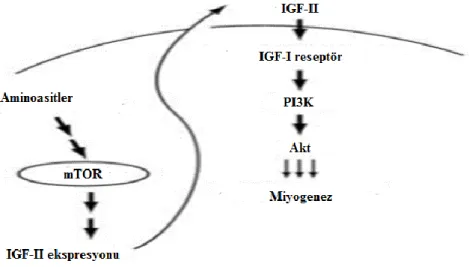
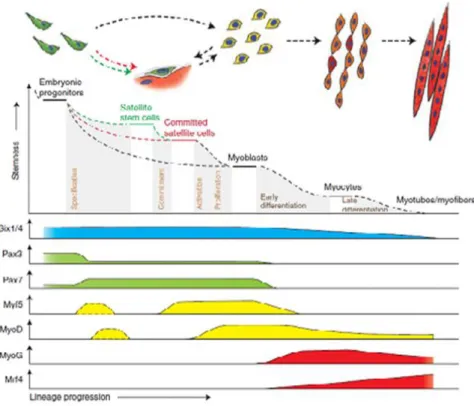
Farelerde resveratrol ile beslenmenin myogenin ve mtor düzeyleri üzerine etkisi
Tam metin
Şekil





Outline
Benzer Belgeler
Sonlu eleman modelinden elde edilen dinamik parametreler Sonlu Eleman Modeli Güncelleme tekniği ile deneysel yolla elde edilen dinamik parametrelere yakınsayıncaya kadar
Dick (1996) tarafından Stigler’in görüşleri ve Webb-Pomere Kartelleri baz alınarak yapılan çalışmada, yoğunlaşmış piyasalara ürün satan kartellerin diğerlerine göre
Özet - 6u çalışmada toprak materyalinin su ve i·üzgarla taşınması olarak tanımlanan erozyonun, özellikle yağışların etkisiyle Adapazarı topraklarına
Yarışmaya katılmak için yazım kurallarına uygun proje öze- ti hazırlanarak en geç 7 Haziran 2013 tarihine kadar Makina Mühendisleri Odası İstanbul Şubesi’ne elden, posta veya
Granger (1969; 424- 428) proposed a method known as the “Granger causality test” and many studies are done belonging to determining the causality relationships between energy
• Karotenoitler, planktonlar, algler, bitkiler ve bazı mikroorganizmalar tarafından doğal
Fa kat söz ko nu su ayet, bü tün va tan daş la rın eşit hak lar ve so rum lu luk la rı nın tes pi tin de dinî iliş ki - ler den zi ya de va tan daş lı ğın esas alın dı
MTT analizi sonucunda, uygulanan dilüsyon aralığında ve 48, 72 ve 96 saat inkübasyon sürelerinde çözücü miktarının sıçan gingival fibroblast canlılığı
