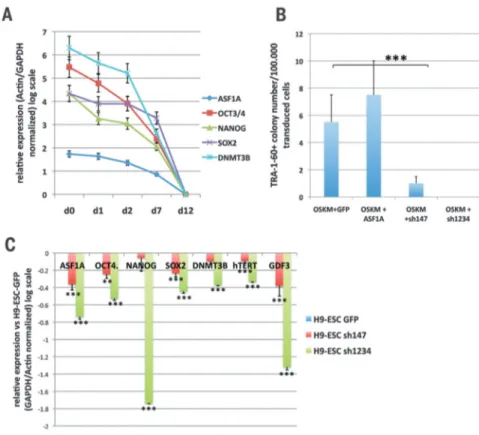
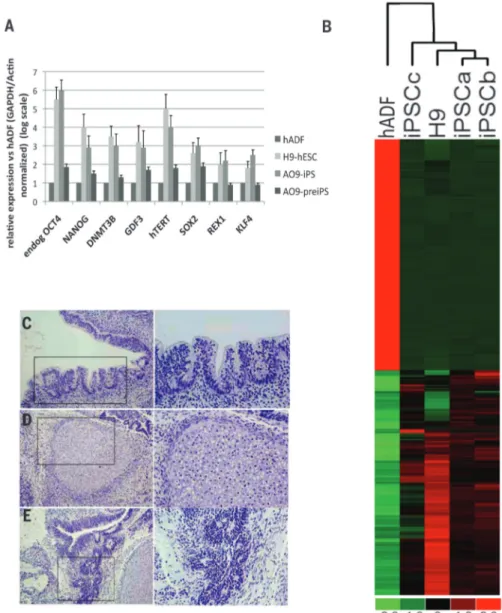
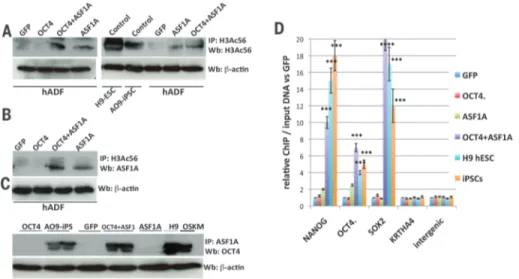
Cell reprogramming. Histone chaperone ASF1A is required for maintenance of pluripotency and cellular reprogramming
Tam metin
Şekil
Benzer Belgeler
Resim pazara dönüştüğünde, resim alıcı (koleksiyoncu, diyemiyorum, çünkü koleksiyoncu bilinçli bir alıcıdır.) resim yaratıcısını etkilediği dönemde, o
These results therefore suggest that the single G sequence of the human CYP21 gene is crucial for the expression of its basal transcription activity, and this may be influenced by
A chemical kinetic-type model for the construction of the molecular clusters of each polymorph is formulated that accounts for adsorption at a heterogeneous site, nucleation,
Sağlık İşletmelerinde Kalite ve Algılanan Hizmet Kalitesinin Ölçülmesi, DEÜ Sosyal Bilimler Enstitüsü Yayınlanmamış Yüksek Lisans Tezi, 2001, İzmir, s.105.. 14
Avrupa Dermatoveneroloji Konseyi EBDV dışında dermatoveneroloji eğitimine yönelik çalışması olan 3 diğer kuruluş Avrupa Dermatovenerıoloji Akademisi (European Academy
Operatör Tercihleri ile Akıllı Telefon Marka Tercihlerinin İki Boyutlu Grafiği Grafik 6’dayüksek gelirli Y kuşağı ile akıllı telefona 5 yıl ve üzerinde bir sürede sahip
Sonuç olarak yapılan çalışma ile ülkemizde veteriner kedi ve köpek kliniklerinde beşeri ilaç kullanımının oldukça yaygın olduğu, veteriner onaylı
Bu çalışmada yeşil davranışların yayınlaşması için önemli olduğu düşünülen yeşil dönüştürücü liderlik ele alınmış ve yeşil dönüştürücü liderliğin
