Investigation of spontaneous differentiation of neural stem cells on synthetic scaffolds
Tam metin
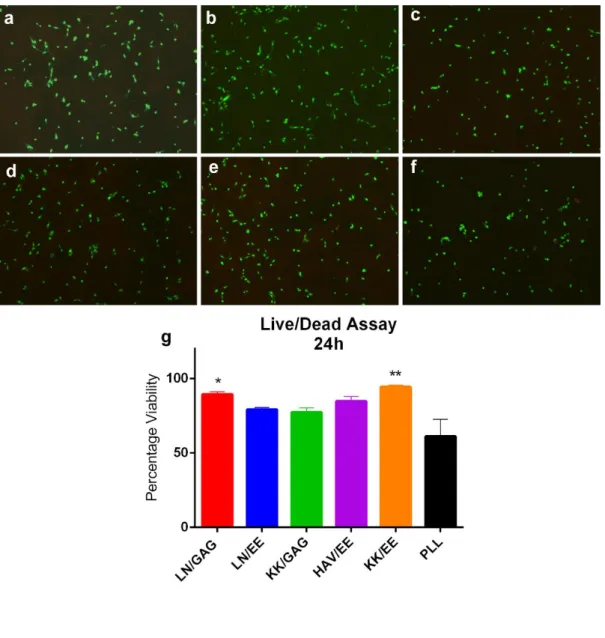
Şekil
![Figure 2 Top down and bottom up approaches of nanomaterial synthesis. Reprinted from ref [2] with permission](https://thumb-eu.123doks.com/thumbv2/9libnet/5844497.119818/20.892.195.772.300.586/figure-approaches-nanomaterial-synthesis-reprinted-ref-permission.webp)
![Figure 3 Self assembling monomers forming various secondary structures [1]. Reprinted with permission from Nature Publishing Group](https://thumb-eu.123doks.com/thumbv2/9libnet/5844497.119818/22.892.188.781.216.825/figure-assembling-monomers-secondary-structures-reprinted-permission-publishing.webp)
![Figure 5 Directing the NSC fate by ECM molecules and other exogenous factors in vivo and in vitro [3]](https://thumb-eu.123doks.com/thumbv2/9libnet/5844497.119818/35.892.180.778.680.1009/figure-directing-nsc-fate-molecules-exogenous-factors-vitro.webp)
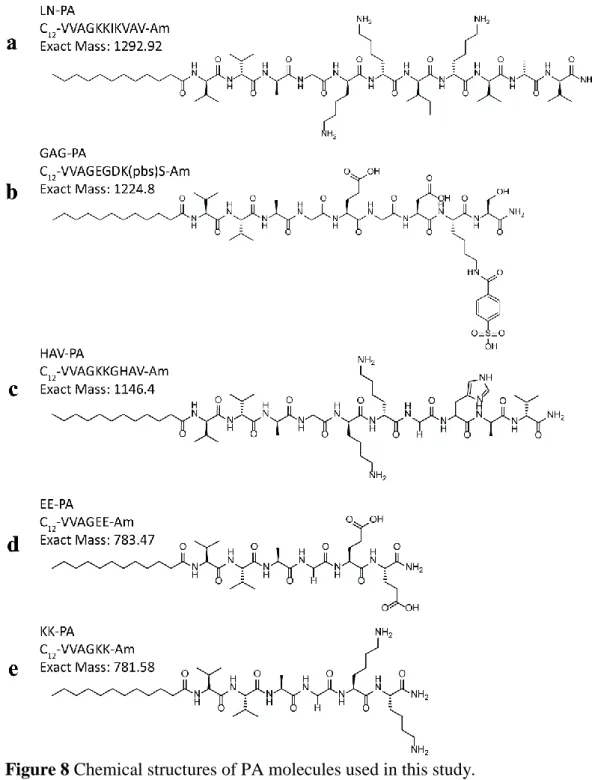
![Figure 9 The LC-MS of the positively charged PAs; (a) LN-PA; [M+H]+](https://thumb-eu.123doks.com/thumbv2/9libnet/5844497.119818/46.892.178.789.287.965/figure-lc-ms-positively-charged-pas-ln-pa.webp)
![Figure 10 The LC-MS of the negatively charged PAs. (a) GAG-PA; [M-H] - (calculated): 1225.59, [M-H] - (observed):1224.61, [M- 2H] -2 /2 (calculated): 612.29, [M-2H] -2 /2 (observed): 611.81, (b) EE; [M-H] - (calculated) = 782.47, [M](https://thumb-eu.123doks.com/thumbv2/9libnet/5844497.119818/47.892.185.791.298.750/figure-negatively-charged-calculated-observed-calculated-observed-calculated.webp)
Benzer Belgeler
Mumun sevgili, pervânenin âfl›k oldu¤undan hareketle beyitten âfl›k olunacak sevgilinin, birçok âfl›¤› bir an- da küle çevirecek bir atefl gibi olmas› ge-
3) Bir kalem, bir eldiven ve bir de kalem traş aldım. Kasaya 50TL verdim. 4) Bir şapka ve bir çift ayakkabı aldım. Kasaya 100TL verdim. Kaç TL para üstü almalıyım?.... 2)
2 A(i) shows FEG-SEM images of the 5 wt.% PCL nano fibrous scaffold at low magnification (500×) and (ii) shows the BC/PCL (50:50 wt.%) nanofibrous scaffold at a much
We show that the cover induced by Strong Nash Equilibrium of the hyper-link formation game for any convex value function and the player based flexible cover allocation rule belongs
Regarding genetic analyses, the GENUS sample collection is best suited for characterizing SNPs, polygenic factors, and pathways identi fied by GWAS, such as the PGC GWAS mega-analyses
“Teknoloji,” diye yazar Wyn Jones (1999: 139), “toplum için bir dizi seçeneğe kapı açar ve seçilmiş opsiyonlar, kısmen o toplum içerisindeki güç
Amongst them liver cell transplantation (LCT) has been receiving great attention in recent years due to the improvement on the isolation and expansion of hepatocyte
Araştırma evreni Düzce İl Merkezi’nde sokakta çalışan çocuklardan ve onların ailelerinden oluşmaktadır. Düzce İl Em- niyet Müdürlüğü Çocuk Şubesi’nden alı-
