i
EGE ÜNİVERSİTESİ TIP FAKÜLTESİ
SPOR HEKİMLİĞİ ANABİLİM DALI
FUTBOLCULARDA VE SEDANTER BİREYLERDE
ACTN3 R577X GEN POLİMORFİZM DAĞILIMI İLE
TELOMERAZ AKTİVİTESİ ARASINDAKİ İLİŞKİ
TIPTA UZMANLIK TEZİ
Hazırlayan
Dr. Fatma Ebru KOKU
Danışman
Prof. Dr. S. Oğuz KARAMIZRAK
iii
EGE ÜNİVERSİTESİ TIP FAKÜLTESİ
SPOR HEKİMLİĞİ ANABİLİM DALI
FUTBOLCULARDA VE SEDANTER BİREYLERDE
ACTN3 R577X GEN POLİMORFİZM DAĞILIMI İLE
TELOMERAZ AKTİVİTESİ ARASINDAKİ İLİŞKİ
TIPTA UZMANLIK TEZİ
Hazırlayan
Dr. Fatma Ebru KOKU
Danışman
Prof. Dr. S. Oğuz KARAMIZRAK
vi
ÖNSÖZ
Uzmanlık eğitimim süresince benden bilgi, tecrübe ve sabrını esirgemeyen tez danışmanım Prof. Dr. S. Oğuz Karamızrak’a, hayatla ve spor hekimliği ile ilgili tecrübelerini cömertçe benimle paylaşan ve yol gösteren Prof. Dr. Çetin İşlegen’e, yetişmemde büyük katkıları olan değerli hocalarım Prof. Dr. K. Ahmet Ertat’a, Prof. Dr. Cengizhan Özgürbüz’e, Prof. Dr. Metin Ergün‘e ve Yrd. Doç. Dr. Seçkin Şenışık’a teşekkürlerimi sunuyorum.
Bu tez çalışmasına fikirleri ve emekleri ile büyük katkıda bulunan Tıbbi Genetik Bilim Dalı Öğretim Üyesi Prof. Dr. M. Özgür Çoğulu’ya, Uzm. Dr. Burak Durmaz’a, Dr. Hasan Taşlıdere’ye, tezimin her aşamasında birlikte çalıştığımız, ve spor hekimliğinde uzmanlık eğitimime başladığım ilk günden beri bana her konuda destek olan Dr. Aynur Sevgi Arslan’a, verilerin toplanması aşamasında benden yardımlarını esirgemeyen Dr. Abdülaziz Türksoylu’ya, Dr. Yavuz Lima’ya, Sayın Burak Aydoğan’a ve tüm Spor Hekimliği Anabilim Dalı personeline teşekkürü bir borç bilirim.
Sevgisi ve sabrıyla beni bugünlere getiren, bana her konuda destek olan canım annem Emine Koku’ya ve tüm aileme teşekkür ediyorum.
KASIM 2014 Fatma Ebru Koku İZMİR
vii
İÇİNDEKİLER
ÖNSÖZ ... vi İÇİNDEKİLER ... vii ŞEKİLLER LİSTESİ ... ix RESİMLER LİSTESİ ... ix TABLOLAR LİSTESİ ... x KISALTMALAR ... xi 1. GİRİŞ ve AMAÇ ...1 2. GENEL BİLGİLER...3 2.1 α-Aktinin-3 ...32.1.1 α-Aktinin-3 Polimorfizminin Toplumda ve Sporcularda Yaygınlığı...4
2.1.2 ACTN3 Genotipinin Sportif Performans ile İlişkisi ...9
2.2 Sürat ve Dayanıklılık Performansı ... 12
2.2.1 Kas Lifi Tipleri ... 12
2.2.2 Enerji Sistemleri ... 15
2.2.3 Anaerobik güç ... 16
2.2.4 Aerobik Güç / Maksimal Oksijen Kullanımı ... 17
2.2.5. Maksimal Oksijen Kullanımına Etki Eden Faktörler ... 20
2.2.6 Anaerobik Eşik ... 22
2.2.7 Maksimal Oksijen Kullanım Kapasitesi Ölçümü ... 23
2.2.8 Futbolda Dayanıklılık ve Sürat Performansı ... 24
2.3 Telomeraz Enzimi ... 24 2.3.1 Telomer ... 24 2.3.1.1 Telomerin Yapısı ... 25 2.3.2 Telomeraz ... 25 2.3.2.1 Telomerazın Yapısı ... 26 3. GEREÇ ve YÖNTEM ... 28 3.1 Araştırmanın Tipi ... 28
3.2 Araştırmanın Yeri ve Zamanı ... 28
3.3 Denekler ... 28
3.3.1 Deneklerin Seçilmesi ... 29
viii
3.3.3 Çalışmadan Çıkarılma Kriterleri ... 29
3.3.4 Çalışmanın Örneklemi ... 29
3.4 Sorgulama ve Fizik Muayene... 30
3.5 Ölçümler ... 30
3.5.1 Boy Uzunluğu Ölçümü ... 30
3.5.2 Subkutan Yağ Ölçümü... 30
3.5.3 Biyoelektriksel Empedans ile Vücut Ağırlığı ve Vücut Yağ Oranı Ölçümü... 31
3.5.4 Durarak Dikey Sıçrama ve Durarak Uzun Atlama Mesafesi Ölçümleri ... 31
3.5.5 Maksimal Oksijen Kullanımı (Maks VO₂) ... 32
3.5.6. Solunumsal Eşik Hesaplanması ... 33
3.5.7 Gen Analizi ve Telomeraz Aktivasyonu Ölçümleri ... 34
a. Örneklerin Alınması ve Saklanması ... 34
b. DNA İzolasyonu ... 34
c. “Real Time” PCR Aşaması ... 35
d. ACTN3 (rs1815739) Gen Polimorfizminin Çalışılması ... 35
e. Veri Analizi ... 36
f. RNA İzolasyon Protokolü ... 36
3.6 İstatistiksel Analiz ... 39
4. BULGULAR ... 40
4.1 Futbolcu ve Kontrol Gruplarının Fiziksel Özellikleri ... 40
4.2 Telomeraz Analizi ... 40
4.3 Futbolcu ve Kontrol Gruplarının ACTN3 R577X Gen Polimorfizmi Dağılımları ... 40
4.4 Futbolcu ve Kontrol Gruplarının Vücut Kompozisyonu Ölçümlerinin Kıyaslaması ... 41
4.5 Futbolcu ve Kontrol Gruplarının Kardiyorespiratuvar Testlerinin Kıyaslaması ... 42
4.6 Futbolcu ve Kontrol Gruplarının Sıçrama Testlerinin Kıyaslaması ... 44
4.7 Futbolcu ve Kontrol Gruplarının Vücut Kompozisyonu ile Performans Testlerinin Korrelasyonu ... 45 5. TARTIŞMA ... 49 6. SONUÇLAR ... 54 7. ÖZET... 55 KAYNAKLAR ... 59 EKLER ... 66
ix
ŞEKİLLER LİSTESİ
Şekil 1: α-Aktininin sarkomerdeki yerleşimi ve α-aktininin fonksiyonel yapısı ...4
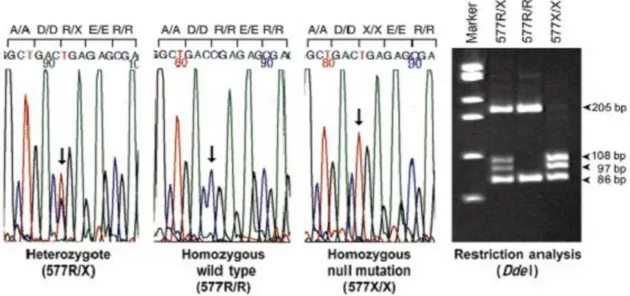
Şekil 2: ACTN3 restriksiyon endonükleaz analizi ve jel elektroforezi ...5
Şekil 3: Üst düzey dayanıklılık ve sürat sporcularında ACTN3 R577X genotip dağılımı ...6
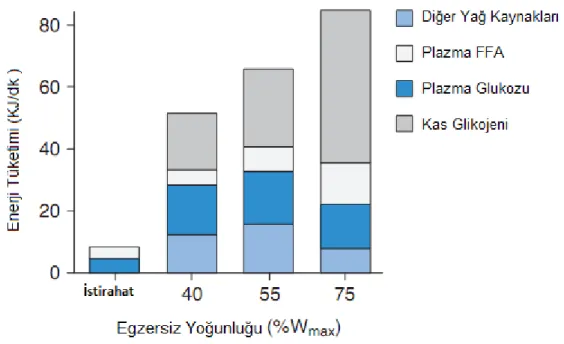
Şekil 4: Egzersizde enerji kaynakları ...16
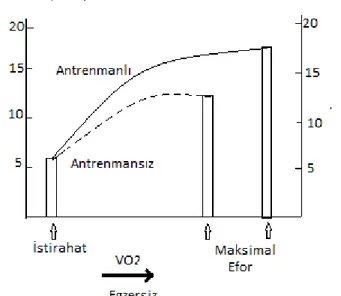
Şekil 5: Antrenmanlı ve antrenmansız bireylerde egzersizin a-v O₂ farkına etkisi: Antrenmanla kas kandan daha fazla oksijen alır ve kullanır ...20
Şekil 6: Artan iş yükünde tüketilen oksijen hacmi(VO₂) ve üretilen karbondioksit hacmi (VCO₂), VO₂(○)’de lineer artış VCO₂(●)’de bifazik artış. ...23
Şekil 7: T-loop ve D-loop yapısı ...25
Şekil 8: Telomeraz bileşenleri ...26
Şekil 9: Yarı açık el yapısı ...26
Şekil 10: Dikey sıçrama ölçümü ...32
Şekil 11: ACTN3 örnek görüntüsü ...36
Şekil 12: Amplifikasyon eğrisi ...39
RESİMLER LİSTESİ
Resim 1: Holtain Skinfold Caliper ...30x
TABLOLAR LİSTESİ
Tablo 1: Farklı spor dallarında ACTN3 R577X polimorfizm dağılımı ...8
Tablo 2: İnsan kasında lif tipi özellikleri ... 14
Tablo 3: Antrenmanlı ve antrenmansız bireylerde oksijen taşıma sisteminin bileşenleri ... 19
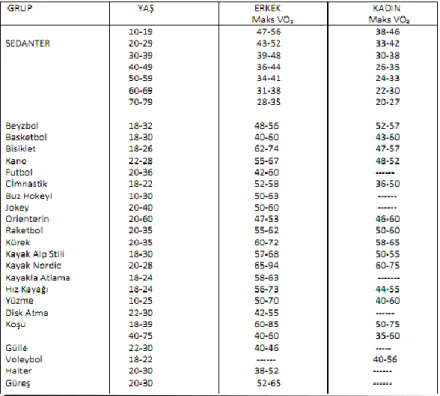
Tablo 4: Farklı yaş grupları ve spor dallarında Maks VO₂ değerleri ... 21
Tablo 5: Roche LightCycler® FastStart DNA Master HybProbe kullanımı ... 35
Tablo 6: Cihaz protokolü... 38
Tablo 7: Kontrol ve futbolcu gruplarının yaş, boy ve VA parametreleri ... 40
Tablo 8: Futbolcu ve kontrol gruplarının ACTN3 R577X gen polimorfizmi dağılımları ... 41
Tablo 9: Futbolcu ve kontrol gruplarının vücut kompozisyonu ölçümleri ... 41
Tablo 10: Futbolcu ve kontrol gruplarının ACTN3 gen polimorfizmi dağılımına göre vücut kompozisyonu ölçümleri ... 42
Tablo 11: Futbolcu ve kontrol gruplarının ACTN3 gen polimorfizmi dağılımına göre skinfold ölçümleri ... 43
Tablo 12: Futbolcu ve kontrol gruplarının kardiyorespiratuvar testlerinin kıyaslaması ... 43
Tablo 13: Futbolcu ve kontrol gruplarının ACTN3 gen polimorfizmi dağılımına göre kardiyorespiratuvar test sonuçları ... 44
Tablo 14: Futbolcu ve kontrol gruplarının sıçrama testlerinin kıyaslaması ... 44
Tablo 15: Futbolcu ve kontrol gruplarının ACTN3 gen polimorfizmi dağılımına göre sıçrama testleri sonuçları ... 45
Tablo 16: Kontrol grubunun vücut kompozisyonu ve performans testlerinin korrelasyonu ... 46
xi
KISALTMALAR
a-v O₂ farkı: Arterio venöz oksijen farkı EBS: Eller belde dikey sıçrama
hTERT: Human telomeraz revers transkriptaz KO fare: Knockout fare
KVH: Kardiyovasküler hastalıklar Maks VO₂: Maksimal oksijen kullanımı MHC: Miyozin ağır zincir
NADH-TR: NADH tetrazolium redüktaz PCR: Polimeraz zincir reaksiyonu PCr: Fosfokreatin
Q: Kalp debisi
RER: Solunum değişim oranı SDH: Süksinat dehidrogenaz
SS: Squat sıçrama
TRAP: Telomere repeat amplification protocol VKİ: Vücut kitle indeksi
VYO: Vücut yağ oranı VA: Vücut ağırlığı VO₂: Oksijen hacmi VCO₂: Karbondioksit hacmi
1
1.
GİRİŞ ve AMAÇ
Sporda performansı etkileyen faktörler, spor bilimleri ve spor hekimliği bilim dallarının önemli araştırma konularıdır. Sportif performansta ya da yetenekte genetik etki ise uzun yıllardır incelenmektedir.
Otozomal genler, mitokondriyal DNA ve Y kromozomunda yer alan pek çok gen sportif performans ile ilişkilendirilmiştir. ACTN3 R577X gen polimorfizmide sıkça incelenmiş ve performans üzerine etkisi ile ilgili farklı sonuçlara ulaşılmıştır. ACTN3 geni, Z çizgisinin temel yapı proteini olan aktin bağlayıcı protein ailesinin üyesi α-aktinin-3’ ü kodlar. ACTN3 ekspresyonu sadece tip 2 (hızlı glikolitik) kas lifileri ile sınırlandırılmıştır ACTN3 R577X polimorfizmi ise 16. ekzonda 1747 pozisyonunda meydana gelen C→T transversiyonu sonucunda 577. pozisyondaki arjinin (R) amino asidini oluşturan kodonun stop kodona (X) dönüşmesidir. Eğer X alleli homozigot (XX genotipi) ise, α-aktinin-3 hiç sentezlenmez (56). Literatürde sürat ve güç ile ilgili spor dallarındaki sporcularda XX genotip sıklığının düşük olduğunu gösteren çalışmalar mevcuttur. Dayanıklılık sporcularında ise XX genotip sıklığının artışı net olarak kanıtlanamamıştır (83).
Telomer ise ökaryotik organizmalarda kromozomların uçlarında yalıtımı sağlamak için yer alan, kodlanmayan özelleşmiş DNA tekrar dizilerinden (memelilerde TTAGGG) oluşan heterokromatik bölgelerdir. Telomeraz enzimi, yapısında bulunan RNA’sını kalıp olarak kullanarak telomerleri uzatan bir revers transkriptazdır (1). Sedanter yaşam tarzı telomer dinamiklerini olumsuz etkiler. Son yıllarda yapılan çalışmalarda, fiziksel olarak aktif bireylerin telomer boyunun, inaktif bireylere göre daha uzun olduğu gösterilmiştir (14,43).
Bu tez çalışmasının amacı; futbolcularda ve sedanter üniversite öğrencilerinde ACTN3 R577X polimorfizm dağılımı ile telomeraz aktivitesi arasındaki ilişkiyi belirlemek ve egzersizin telomeraz aktivitesi üzerindeki etkisini araştırmaktır. Ayrıca; spor yapan RR genotipindeki bireylerde anaerobik güç test sonuçlarının spor yapan XX genotipine sahip bireylere göre daha iyi olup olmadığını ve spor yapan XX genotipine sahip bireylerin Maks VO2 değerlerinin, spor yapan RR genotipine sahip bireylere göre daha iyi olup olmadığını incelemektir.
Çalışmanın hipotezleri;
2
XX genotipine sahip bireylerin telomeraz aktivitesi, RX ve RR genotipine sahip bireylere göre daha yüksektir.
XX genotipine sahip futbolcularda telomeraz aktivitesi, XX genotipine sahip sedanter bireylere göre daha yüksektir.
XX genotipine sahip futbolcularda telomeraz aktivitesi, RX ve RR genotipine sahip tüm bireylerden daha yüksektir.
XX genotipine sahip sedanterlerin telomeraz aktivitesi, RX ve RR genotipine sahip sedanterlerden daha yüksektir.
3
2. GENEL BİLGİLER
Spor, günümüzde pek çok insanın ilgilendiği bir uğraş olup kitleler arasında hızla yaygınlaşmaktadır. Spora amatör ya da profesyonel katılım arttıkça sportif performans ve bu performansı etkileyen faktörlerin önemi de artmaktadır. Sporda başarıyı belirleyen faktörler genetik, epigenetik, beslenme, antrenman, motivasyon, ekipman ve diğer çevresel faktörler altında sınıflandırılabilir. Spor ve fiziksel performans genetiği terimini ilk kez çeyrek asır önce Claude Bouchard kullanmış ve bu alanda ilk yayını yapmıştır (11,13). Fiziksel performansta modern genetik dönemi ise 1990’ların sonunda “İnsan Genom Projesi” ile başlamıştır. Genetiğin kuvvet, güç, dayanıklılık, kas lifi tipi, boyutu, kompozisyonu, esneklik, nöromüsküler koordinasyon, karakter ve diğer fenotiplere etkisi büyüktür. Sportif durumun %66’sını genetik belirlerken, kalan kısmı çevresel faktörler belirler (18). Otozomal genler, mitokondriyal DNA ve Y kromozomunda yer alan pek çok gen sportif performans ile ilişkilendirilmiştir. Bu genlerden biri de α-aktinin-3’ü kodlayan ACTN3 genidir.
2.1 α-Aktinin-3
Hareket sistemimizin temelini kaslar oluşturur. İskelet kası birbirinden bağımsız kas liflerinden oluşur. Her kas lifi, birden çok çekirdek içeren uzun silindirik tek bir kas hücresinden oluşmuştur. Kas hücresinin hücre zarına sarkolemma denir. Bu zar sarkoplazma adı verilen hücre plazmasını çevreler.
Her lif sarkoplazma içerisinde asılı halde duran yüzlerce miyofibrilden oluşmuştur. Miyofibriller ise protein yapısındaki ince ve kalın miyofilamentlerden meydana gelirler. Bunlardan ince miyofilament ağırlıklı olarak aktin olmak üzere troponin ve tropomiyozin moleküllerinden, kalın miyofilament ise miyozin moleküllerinden oluşmuştur. İskelet kasına çizgili görünümü veren aktin ve miyozin filamentlerinin dizilişidir. Yalnızca aktin filamentlerinin bulunduğu bölge I (izotropik) bandı adını alır ve ışık mikroskopunda açık renk görülür. Aktin ve miyozin filamentlerinin birlikte yer aldığı kısımlar daha koyu renk görülürler ve A (anizotropik) bandı olarak isimlendirilirler. A bandının ortasında aktinin ulaşamadığı ve yalnızca miyozinden oluşan bir alan vardır ve H (Hensen diski) bölgesi olarak adlandırılır. I bandının ortasında dikey olarak uzanan ve kas lifinin içindeki miyofibrilleri birbirine bağlayan Z (Zwischenscheibe) çizgisi bulunur. İki Z çizgisi arasında kalan bu bölgeye sarkomer adı verilir. Sarkomer kasılabilen en küçük birimdir (27).
4 α-Aktininler ise hücre iskeletini oluşturan ve düzenleyen aktin bağlayıcı protein ailesinin (distrofin, spektrin, filamin, fimbrin) bir üyesidir. α-Aktininler iskelet kasında Z çizgisinin temel yapı komponentidir. Çeşitli fonksiyonel rolleri olan yapı, sinyal ve metabolik proteinlerin, protein-protein etkileşimleri için çok değerlikli bir platform sağlar (19). Bu etkileşimle yapı ve sinyal proteinlerinin Z çizgisi ile ilişkisini sağlar. Özellikle iskelet aktinini ve sarkomerik α-aktininler (-2 ve -3) miyofibriller dizilişin statik işlevini korur ve miyofibril kontraksiyonunu düzenler (47). Yapısında N-terminal aktin bağlanma bölgesi, merkezi rod ve kalsiyumun bağlandığı “EF hand” içeren C-terminal bölgesi olmak üzere üç fonksiyonel bölge vardır (9). N-terminal uç ile iskelet α-aktinine ve C-terminal uç ile diğer Z çizgisi proteinlerine (myotilin v.b.) bağlanarak sarkomerin üç boyutlu örgü yapısını korumasına katkıda bulunur. Sarkomerik α-aktininler distrofin ve integrin adezyon sistemini de bağlayarak kas hücre bütünlüğünün korumasına katkı sağlarlar (33).
Memelilerde dört çeşit α-aktinin izoformu vardır ve farklı hücrelerde benzer işlevleri üstlenirler (7,38,41,74). ACTN2 ve ACTN3 tarafından kodlanan α-aktinin-2 ve α-aktinin-3 iskelet kası izoformlarıdır. ACTN2 ve ACTN3’ün amino asit sıralaması %91 özdeş ve %86 benzerdir. ACTN2 tüm iskelet kas lifleri (hızlı ve yavaş lifler), kalp kası ve beyinde eksprese edilirken, ACTN3 ekspresyonu sadece tip 2 (hızlı glikolitik) kas lifileri ile sınırlandırılmıştır. Ancak az miktarda beyinde de eksprese edilir (7).
Şekil 1: α-Aktininin sarkomerdeki yerleşimi ve α-aktininin fonksiyonel yapısı (47)
2.1.1 α-Aktinin-3 Polimorfizminin Toplumda ve Sporcularda Yaygınlığı
ACTN3 R577X polimorfizmi, 16. ekzonda 1747 pozisyonunda meydana gelen C→T transversiyonu sonucunda 577. pozisyondaki arjinin (R) amino asidini oluşturan kodonun stop kodona (X) dönüşmesidir. Eğer X alleli homozigot (XX genotipi) ise, α-aktinin-3 hiç
5 sentezlenmez (56). XX genotip sıkılığı farklı popülasyonlarda değişmektedir. Afrikalılarda ~%6, Afrika Bantu kabilesinde ~%1, Avrupalılarda ~%18, Asyalılarda ~%25 sıklıkta saptanmıştır. Dünya genelinde yaklaşık 1 milyar kişide α-aktinin-3 eksikliği olduğu tahmin edilmektedir. Bu yaygın eksikliğe rağmen hızlı glikolitik liflerin ekpresyonunda kısıtlılık görülmez, bunun nedeni α-aktinin-2 tarafından eksikliğin giderilmesidir (51).
Şekil 2: ACTN3 restriksiyon endonükleaz analizi ve jel elektroforezi (56)
ACTN3 geni evrim süreci boyunca son derece korunmuştur. İnsan, fare ve tavuk ACTN genleri incelendiğinde, insanda ACTN2 ve ACTN3 geninin yaklaşık 300 milyon yıl önce ayrıştığı ve onlar tarafından kodlanan proteinlerin de ayrışmadan beri oldukça yavaş evrimleştiği gösterilmiştir. ACTN2 ve ACTN3 insanlarda ve farelerde embriyonik gelişim boyunca farklı eksprese edilmektedir ve ACTN2 ekspresyonu ACTN3 ile tam olarak çakışmaz. ACTN3 fare ve insanlarda hızlı liflerde baskındır.
Bireyin sürat sporlarında mı, yoksa dayanıklılık sporlarında mı başarı yakalayacağının evrimsel olarak belirlendiğini söyleyebiliriz. Yüksek hızda ve kapasitede kuvvet üreten hızlı glikolitik kas liflerinin antrenmana yanıtında genetik etkinin önemi büyüktür (70). ACTN3 genotipi iskelet kası performansını etkiler ve α-aktinin-3 varlığı ya da yokluğu hızlı kas lifleri tarafından gerçekleştirilen sürat ve güç faaliyetlerini etkileyebilir.
Druzhevskaya, 486 Rus sürat/güç sporcusu ve 1197 Rus sedanter birey ile yaptığı çalışmada kontrol grubunun genotip dağılımını RR %36.8, RX %49.0, XX %14.2 olarak saptamıştır. Bu dağılım Kafkas ırkındaki genotip dağılımına benzerdir. Sporcu grubunda ise genotip dağılımını RR %39.7, RX %53.9, XX %6.4 olarak saptamıştır. Sporcu grubundaki XX genotip oranı kontrol grubuna göre oldukça düşüktür (p<0.0001) (20). Yang, 107 sürat sporcusu, 194 dayanıklılık sporcusu ve 436 sedanter birey ile yaptığı çalışmada sürat
6 sporcularında XX genotipini %6, kontrol grubunda %18 saptamıştır ve bu fark istatistiksel olarak anlamlıdır (p<0.01). Olimpiyat düzeyinde yarışan kadın süratçilerde ise hiç XX genotipine rastlamamıştır (Şekil 3). Kadın dayanıklılık sporcularında ise XX genotipini (%29), kontrol grubundan (%20) yüksek saptamıştır (p<0.05) (84).
Şekil 3: Üst düzey dayanıklılık ve sürat sporcularında ACTN3 R577X genotip dağılımı (84)
Ahmetov ise 456 dayanıklılık sporcusu grubunu 1211 sedanter kontrol grubu ile karşılaştırdığı çalışmasında, XX genotipini dayanıklılık sporcularında (%5.7) kontrol grubuna (%14.5) göre düşük saptamıştır (p<0.0001) (2). Bahsedilen bu iki çalışmanın dayanıklılık sporcularındaki sonuçları birbiri ile çelişmektedir. Eynon, 305 dayanıklılık sporcusu, 378 sürat/güç sporcusu, 205 takım sporcusu ve 568 sedanter birey ile yaptığı araştırmada; takım sporu yapanların genotip dağılımını, dayanıklılık sporcuları ve kontrol gruplarında benzer olarak saptamıştır. Takım sporu yapanların RR genotip oranı, sürat/güç sporcularının RR genotip oranına göre düşük bulunmuştur (p=0.045) (23).
Roth, vücut geliştirme ve ağırlık kaldırma sporu ile ilgilenen 75 sporcu (52 beyaz, 23 zenci) ve 876 sedanter (668 beyaz, 208 zenci) birey ile yaptığı çalışmada, sporcularda XX genotipini (%6.7), kontrol grubuna (%16.3) göre oldukça düşük saptamıştır (p=0.005). Sadece beyaz ırka bakıldığında, XX genotipi sporcularda %9.7, beyaz ırk kontrol grubunda %19.9 (p=0.018) oranında saptanmıştır. Zenci sporcularda ise hiç XX genotipine rastlanmamıştır (64).
Papadimitriou, 34 kısa mesafe koşucusunu 181 sedanter birey ile karşılaştırdığı çalışmasında, sporcularda genotip dağılımını RR %73.5, RX %17.7, XX %8.8; kontrol grubunda ise RR %26.0, RX %55.8, XX %18.2 olarak saptamıştır. Sporcu grubunda XX oranını kontrol grubuna göre oldukça düşük (p<0.001), RR oranını ise yüksek (p=0.0001) saptamıştır (57). Niemi, Finlandiyalı uzun mesafe koşucularında (n=40) genotip dağılımını, kontrol grubunun (n=120) genotip dağılımı ile benzer bulmuş; üst düzey kısa mesafe koşucularında ise (n=23) hiç XX genotipine rastlamamıştır (p=0.03) (55).
7 Yang; Kenya, Nijerya ve Etyopyalı sporcu ve sedanter bireylerden oluşan Afrika polülasyonunu ACTN3 R577X gen polimorfizm dağılımı açısından araştırmıştır. Nijeryalı sürat/güç sporcularında ve kontrol grubunda hiç XX genotipine rastlamamıştır. Etyopyalı kontrol grubunda XX genotipini %12 oranında saptamıştır. Beklenenin aksine, dayanıklılık sporcularında XX genotipi sıklığı (%6) istatistiksel fark yaratmasa da kontrol grubuna göre düşüktür. Kenyalı dayanıklılık sporcuları ve kontrol grubunda ise XX genotipini %1 oranında belirlemiştir (85).
Farklı branşlar ele alındığında; Ruiz’in voleybolcularda (n=66), Paparini’nin İtalyan kürekçilerde (n=42), Saunders’in triatloncularda (n=457), Lucia’nın bisikletçi (n=50) ve uzun mesafe koşucularında (n=52), Muniesa’nın İspanyol kürekçilerde (n=39), Garatachea’nın basketbolcularda (n=100) ve Bell’in ragbi sporcularında (n=102) yaptıkları çalışmalarda; sporculardaki ACTN3 R577X polimorfizm dağılımları ile kontrol gruplarındaki polimorfizm dağılımları benzer bulunmuştur (18,29,45,54,58,65,68).
Tüm bu çalışmalar değerlendirildiğinde; üst düzey sürat/güç sporcularında XX genotip sıklığının düşük olduğu söylenebilir, ancak ACTN3 geninin dayanıklılık performansı üzerine etkisi ile ilgili sonuçlar çelişkilidir.
Futbolcularda yapılan çalışmalar incelendiğinde; Santiago 60 üst düzey futbolcuyu, 123 sedanter ve 52 dayanıklılık sporcusu ile karşılaştırmıştır. Genotip dağılımını futbolcularda RR %48, RX %37, XX %15; dayanıklılık sporcularında RR %26.5, RX %52.0, XX %21.5; sedanterlerde ise RR %29, RX %54, XX %17 olarak saptamıştır. Futbolcularda RR sıklığını, kontrol grubu ve dayanıklılık sporcu grubuna göre anlamlı olarak yüksek, RX sıklığını ise düşük belirlemiştir (p= 0.041). Araştırmacı bu dağılıma göre üst düzey futbolcuların genotipik olarak sürate yatkın olduğu sonucuna varmıştır (66).
Pimenta, 37 Brezilyalı futbolcu ile yaptığı çalışmada ACTN3 polimorfizm dağılımını RR %40.5, RX %35.2, XX %24.3 olarak bulmuştur (61). Yine Pimenta 200 üst düzey (birinci lig) Brezilyalı futbolcuyu incelediği çalışmasında ACTN3 polimorfizm dağılımını RR %45, RX %44, XX %11 olarak saptamıştır (62).
Massidda, Serie A (n=46) ve Primavera’da (n=44) oynayan 90 futbolcuda ACTN3 polimorfizm dağılımını RR %31, RX %64, XX %5 olarak (50); Eynon ise 50 üst düzey İspanyol futbolcuda ACTN3 polimorfizm dağılımını RR %46, RX %36, XX %18 olarak saptamıştır (23). Futbolcularda belirlenen bu dağılımlar birbirinden oldukça farklıdır.
8
Tablo 1: Farklı spor dallarında ACTN3 R577X polimorfizm dağılımı
Araştırmacı/Ülke Grup (cins, n) Genotip (%) RR RX XX
*p değeri Ref No Yang/AUS Sürat (E, 72)
Dayanıklılık (E, 122) Kontrol (E, 134) Sürat (K, 35) Dayanıklılık (K, 72) Kontrol (K, 292) 53 39 08* 28 52 20 30 54 16* 43 57 00* 36 35 29 30 50 20* <0.001 <0.01 84
Druzhevskaya/RUS Sürat/güç (E, 363) Kontrol (E, 524) Sürat/güç (K, 123) Kontrol (K, 673) 37.7 55.9 06.4* 36.8 46.8 16.4* 45.5 48 06.5 36.8 50.7 12.5 <0.0001 20 Ruiz/ESP Voleybol (E, 31)
Kontrol (E, 243) Voleybol (K, 35) Kontrol (K, 91) 26.6 64.5 12.9 31.7 48.1 20.2 20.0 65.7 14.3 29.7 57.1 13.2 65 Santiago/ESP Futbol (E, 60)
Kontrol (E, 123)
48.3* 36.7 15.0 28.5* 53.7 17.8
=0.041 66 Lucia/ESP Uzun mesafe koşu (E, 52)
Yol bisikleti (E, 50) Kontrol (E, 123)
25 58 17 28 46 26 28 54 18
45 Roth/USA Vücut geliştirme (mix, 52)
Kontrol (mix, 668) 25.0 65.4 09.6* 32.6 47.5 19.9* =0.018 64 Papadimitriou/GRE Güç (mix, 73) Sürat (mix, 34) Dayanıklılık (mix, 28) Kontrol (mix, 181) 48 35.6 16.4 73.5* 17.7 08.8* 50 25 25 26.0* 55.8 18.2* <0.001 57 Niemi/FIN Uzun mesafe koşu (40)
Sürat (23) Kontrol (120) 50 40 10 48 52 00* 45 46 09* =0.03 55 Paparini/ITA Kürek (E, 42)
Kontrol (E, 102) 21 60 19 31 47 22 58 Muniesa/ESP Kürek (39) Kontrol (123) 33 44 26 28 54 18 54 Garatachea/ESP Basketbol (E, 60)
Kontrol (E, 216) Basketbol (K, 40) Kontrol (K, 67) 20.0 26.0 14.0 32.4 47.7 19.9 42.5 40.0 17.5 29.9 56.7 13.4 29 Yang/ETH Dayanıklılık (mix, 76)
Kontrol (mix, 198)
46 46 08 42 47 11
85 Yang/KEN Dayanıklılık (mix, 284)
Kontrol (mix, 158)
75 24 01 84 15 01
85 Yang/NIG Sürat/Güç (mix, 62)
Kontrol (mix, 60)
87 13 00 83 17 00
85 Bell/UK Ragbi (E, 102)
Kontrol (E, 110)
29 54 17 34 48 18
18 Ahmetov/RUS Dayanıklılık (E, 293)
Kontrol (E, 532) Dayanıklılık (K, 163) Kontrol (K, 679) 40 53 07* 36 47 17* 37 59 04* 37 50 13* <0.01 <0.01 2
9
2.1.2 ACTN3 Genotipinin Sportif Performans ile İlişkisi
R577X sportif durum ve insan kas performansı ile ilişkilidir, α-aktinin-3 eksikliği hızlı kasılan liflerin fonksiyonunu etkiler, kas metabolizmasında aerobik yolakları aktive edip dayanıklılığı arttırır. Literatürde 577XX genotipinin sürat ve güç ile ilgili spor dallarındaki sporcularda düşük olduğunu gösteren çalışmalar mevcuttur. Dayanıklılık sporcularında ise XX genotip sıklığının artışı net olarak kanıtlanamamıştır (83). Ayrıca sedanterlerde yapılan iki çalışmada da azalmış kas gücü (15) ve zayıf sürat performansı (53) ile 577XX genotipi ilişkili bulunmuştur. α-Aktinin-3 eksikliğinin kas yapısı ve fizyolojik özellikler üzerindeki etkisini anlamak için fare modeli incelenebilir. Mac Arthur’un “knockout fare” (KO) modeliyle yaptığı çalışmada, α-aktinin-3 ekspresyonunun azalmasının, kasta aerobik yolakları aktive ederek dayanıklılığı arttırdığı gösterilmiştir. KO (ACTN3-/-) farelerde α-aktinin-3 proteinin hiç sentezlenmediği saptanmıştır. KO fareler ve doğal tip (ACTN3+/+) fareler morfolojik olarak benzerdir. KO (ACTN3-/-) farelerde ışık ve elektron mikroskobunda sarkomerlerin normal yapıda olduğu gösterilmiştir. Hızlı glikolitik (tip 2b) liflerde önemli bir kayıp saptanmamıştır, çünkü hızlı liflerdeki α-aktinin-3 kaybı, α-aktinin-2 ile giderilmektedir.
Knockout (KO) fare çalışmasında α-aktinin-3 eksikliğinin kas metabolizmasında değişikliğe neden olup olmadığı araştırılmıştır. ACTN3-/- farelerin ACTN3+/+ olanlara göre aerobik metabolizmanın iki belirleyicisi olan NADH tetrazolium redüktaz (NADH-TR) ve süksinat dehidrogenaz (SDH) açısından daha yoğun boyandığı gösterilmiştir. Oksidatif enzim aktivitesi artış nedeninin hızlı liflerin kaybı değil, bu liflerin metabolizmasının değişmesi olduğu gösterilmiştir. ACTN3-/- farelerde mitokondri yoğunluğunun arttığı ve mitokondriyal belirleyiciler olan sitokrom c oksidaz ve mitokondriyal porinin yukarı yönlü regüle olduğu saptanmıştır. Glikolitik yolun son metaboliti olan pirüvatın metabolizmasında anahtar rol oynayan iki enzim sitrat sentaz ve laktat dehidrogenaz (LDH) incelenmiştir. Hızlı lifler anaerobik yolda pirüvatı laktat dehidrogenaz ile laktata dönüştürürken, oksidatif yavaş lifler aerobik yolda ilk reaksiyonu sitraz sentaz ile katalizler. ACTN3-/- farelerde LDH aktivitesi ACTN3+/+ olanlara göre %16 düşük (p<0.001), sitrat sentaz aktivitesi %22 yüksek (p<0.05) saptanmıştır. İskelet kasındaki bu metabolizma değişiminin dayanıklılık performansına etkisini göstermek için farelere egzersiz kapasitesi testi yapılmış; fareler artan hızlarda tükeninceye kadar koşturulmuştur. ACTN3-/- farelerin egzersiz kapasitesi %33 daha yüksek saptanmıştır (49).
a. ACTN3’ün Kas Kasılması Üzerine Etkisi
Knock out fareler ve doğal tip farelerin extensor digitorum longus (hızlı kasılan) kasları izole edilmiş ve KO farelerde doğal tipe göre yarı gevşeme zamanının daha uzun olduğu ve yorgunluktan toparlanmanın daha iyi olduğu gösterilmiştir. Bu bulgular ile KO farelerde hızlı
10 kasılan kasların, yavaş kasılan kas liflerinin karakterini kazandıkları ve sonuç olarak doğal tip farelere göre oldukça uzun mesafe koştukları gösterilmiştir (49).
b. ACTN3’ün Kas Kuvveti ve Gücü Üzerine Etkisi
α-Aktinin-3 eksikliğinin sonuçları çok göze çarpmamaktadır, ancak insan ve farelerde kas gücünü azalttığı gösterilmiştir. ACTN3 KO farelerde kavrama gücü doğal tip farelere göre erkeklerde %7.4 (p=0.007), dişilerde %6.0 (p=0.02) oranında düşük saptanmıştır (48). Sedanter insanlarla yapılan bazı çalışmalarda kas gücünün azaldığı gösterilmiştir. XX genotipine sahip kadınlarda RR genotipine göre dirsek fleksör kaslarının izometrik kuvveti düşük saptanmıştır (p<0.05) (15). Genç erkeklerde (n=90, 18-29 yaş) diz ekstansiyon kuvvetinin ölçüldüğü bir çalışmada 300°/s rölatif dinamik kuadriceps gücü XX genotipli katılımcılarda düşük saptanmıştır (p<0.05) (15). Kadınlarda (n=394, 22-90 yaş) yapılan başka bir çalışmada ise diz ekstansörlerinin konsantrik (p=0.019) ve eksantrik (p=0.022) kuvveti XX genotipinde düşük saptanmıştır (80).
Yunanlı adolesan erkeklerin (n=525) katıldığı bir çalışmada ise XX genotipinde olanlar 40 m sürat koşusunu daha uzun sürede tamamlamışlardır (ortalama zaman XX: 6.13 s, RX: 6.00 s, RR: 5.92 s, p=0.003) (53). Genotipin yaşlanma sürecine etkisini inceleyen beş yıl izlemli bir çalışmada ileri yaş kadınlarda (70-79 yaş) XX genotipe sahip olanlar RR genotipine sahip olanlara göre yürüme ve merdiven çıkmada zorluk yaşamaktadır. İleri yaş erkeklerde (70-79 yaş) ise XX genotipine sahip erkekler 400 metreyi daha uzun sürede yürümüşlerdir (17).
Sporcularda ACTN3 R577X polimorfizminin kas gücü üzerine etkisini inceleyen pek çok çalışma bulunmaktadır. Pimenta 200 üst düzey Brezilyalı futbolcuda, squat sıçrama (SS) ve eller belde dikey sıçrama (EBS) testlerinde RR ve RX genotipinde olan futbolcuların, XX genotipinde olan futbolculara göre daha yüksek değerlere ulaştıklarını saptamıştır (p<0.05). Ayrıca sporculara 10m, 20m ve 30m sürat testi uygulamış; 10 metre sürat testinde RR genotipindeki bireylerin XX genotipinde olanlara göre daha süratli (p<0.05) olduklarını; 20 ve 30 m testlerinde ise RR genotipinde olanların RX ve XX genotipinde olanlara göre daha süratli (p<0.05) olduklarını saptamıştır (62).
Massidda 90 üst düzey futbolcuda (Serie A, n=46; Primavera, n=44) sıçrama testleri ile ACTN3’ü de içeren altı farklı genetik polimorfizmin ilişkisini araştırmıştır. Eller belde dikey sıçrama test sonuçları Primavera’da 40.9 ± 4.0 cm, Serie A’da 42.2 ± 4.0 cm olarak ölçülmüştür. Squat sıçrama ise Primavera’da 38.8 ± 3.9 cm, Serie A’da 40.2 ± 4.1 cm olarak belirlenmiştir. Araştırmacı ACTN3, ACE ve BDKRB2 genlerinin dikey sıçramada etkili olduğunu belirtmiştir (50).
11 Bell, futbol gibi bir takım sporu olan ragbi branşında yarışan sporcularda dikey sıçrama testi (EBS) ile ACTN3 genotip ilişkisini incelemiş; ACTN3 genotip dağılımına göre sıçrama testlerinde anlamlı fark belirlememiştir (8).
Ruiz, 66 elit voleybolcuda (31 erkek, 35 kadın) ve aynı yaş grubu 334 sedanter bireyde (243 erkek, 91 kadın) ACTN3 R577X genotip dağılımı ile sıçrama testlerinin ilişkisini araştırmış; ACTN3 genotip dağılımınının her iki grupta benzer olduğunu saptamıştır. Kontrol erkeklerde squat sırçama (SS) 38.5 ± 5.5 cm, eller belde dikey sıçrama (EBS) 40.1 ± 5.8 cm, voleybolcu erkeklerde SS 40.6 ± 6.3 cm, EBS 44.8 ± 7.8 cm olarak ölçülmüştür. SS değerleri kontrol grubunda RR (n=77) genotipinde 38.8 cm, RX (n=117) genotipinde 39.0 cm, XX (n=49) genotipinde ise 39.6 cm; EBS değerleri ise kontrol grubunda RR genotipinde 40.4 cm, RX genotipinde 40.5 cm, XX genotipinde ise 40.0 cm olarak ölçülmüştür. Sporcu grubunda SS değerleri RR (n=14) genotipinde 40.9 cm, RX (n=43) genotipinde 41.0 cm, XX (n=9) genotipinde ise 38.2 cm; EBS değerleri ise RR genotipinde 44.2 cm, RX genotipinde 45.3 cm, XX genotipinde ise 43.2 cm olarak ölçülmüştür. ACTN3 genotipine göre sıçrama testlerinde fark saptanmamıştır (p>0.05) (65).
Garatachea; 100 (60 erkek, 40 kadın) üst düzey basketbolcu ve 283 (216 erkek, 67 kadın) sedanter bireyde ACTN3 R577X genotip dağılımı ile sıçrama testlerinin ilişkisini incelemiştir. Sporcu grubu ve kontrol gruplarının genotip dağılımı bu çalışmada benzer saptanmıştır. Squat sıçrama (SS) değerleri kontrol grubu erkeklerde 38.6 ± 5.5 cm, sporcu grubu erkeklerde 37.4 ± 5.4 cm; eller belde dikey sıçrama (EBS) değerleri kontrol grubu erkeklerde 40.1 ± 5.8 cm, sporcu grubu erkeklerde ise 39.7 ± 5.1 olarak ölçülmüştür. İki grup arasında istatistiksel fark saptanmamıştır. Her iki grupta sırçama testleri genotip dağılımına göre incelendiğinde ise SS düzeyleri kontrol grubunda RR (n=70) genotipinde 38.9 cm, RX (n=103) genotipinde 38.8 cm, XX (n=43) genotipinde ise 37.7 cm olarak; EBS düzeyleri kontrol grubunda RR genotipinde 39.9 cm, RX genotipinde 40.2 cm, XX genotipinde ise 40.0 cm olarak ölçülmüştür. Sporcu grubunda ise SS düzeyleri RR (n=20) genotipinde 38.3 cm, RX (n=26) genotipinde 37.1 cm, XX (n=14) genotipinde 36.2 cm; bu grupta EBS düzeyleri ise RR genotipinde 40.6 cm, RX genotipinde 39.7 cm, XX genotipinde 38.6 cm olarak ölçülmüştür. ACTN3 genotipine göre sıçrama testlerinde anlamlı farklılık saptanmamıştır (p>0.05) (29).
c. Kas Kütlesi, Lif Çapı ve Lif Tipi Oranı Üzerine Etkisi
ACTN3 KO farelerde “dual energy X-ray absorptiometry” (DEXA) ölçümünde, yağsız doku kütlesinde azalma saptanmıştır. KO tipte farelerde hızlı kasılan lifler içeren kas kütlesi de doğal tip farelere göre düşük saptanmıştır (78). İnsanlarda yapılan bir çalışmada da XX genotipindeki kadınlarda, RR ve RX olanlara göre yağsız doku kütlesi (p=0.04) düşük, yağ dokusu (p=0.02) düşük, ayrıca XX genotipinde olanların beden kitle indeksi RR olanlara göre
12 düşük saptanmıştır (p=0.013), erkeklerde ise fark saptanmamıştır (80). Yine postmenapozal kadınlarda yapılan bir çalışmada XX genotipine sahip olanlarda uyluk kas kütlesinin RR genotipine sahip olanlara göre daha az olduğu saptanmıştır (p<0.05) (87). RR (n=22) ve XX (n=22) genotipine sahip genç erkeklerin (18-29 yaş) sağ vastus lateralis kasından alınan biyopsilerin karşılaştırıldığı bir araştırmada ise tip 2X lifler RR genotipinde XX genotipine göre yüksek saptanmıştır (p<0.05) (78).
d. Kas Hasarı Üzerine Etkisi
Brezilyalı futbolculardada yapılan bir çalışmada eksantrik egzersiz sonrasında XX genotipine sahip grupta kas hasarını gösteren kreatin kinaz (CK), α-aktinin ve kortizol düzeyleri, RX ve RR genotipine sahip gruba göre yüksek saptanmıştır. Eksantrik egzersizler tip II liflerde kas hasarına neden olur. Sözü geçen çalışma ile XX genotipine sahip grupta kas hasarını gösteren belirteçlerin yüksek olduğu gösterilmiştir (61). Yine bir başka araştırmada, maksimal eksantrik diz ekstansiyonundan sonra XX genotipindeki katılımcılarda kan CK düzeyleri ve ağrı skorları, RR genotipindeki katılımcılara göre yüksek saptanmıştır (79). Bu çalışmalar doğrultusunda α-aktinin-3’ün hızlı liflerde eksantrik egzersiz sırasında muhtemel koruyucu etkisi olduğu söylenebilir.
2.2 Sürat ve Dayanıklılık Performansı
Sürat ve dayanıklılık performansını anlayabilmek için öncelikle kas lifi tiplerinden, enerji metabolizmasından, aerobik ve anaerobik güçten bahsetmek gerekir.
2.2.1 Kas Lifi Tipleri
Kas lifleri; histokimyasal, biyokimyasal, morfolojik ve fizyolojik özelliklerine göre farklı şekillerde sınıflandırılabilir. Kasların sınıflandırılması ilk kez Ranvier (1873) tarafından yapılmıştır. Kaslar görünümlerine göre kırmızı kaslar ve beyaz kaslar olarak adlandırılmıştır. Günümüzdekine benzer sınıflama ise Peter (1972) tarafından kasılma özelliklerine ve oksidatif kapasitelerine göre Tip I ve Tip II olarak yapılmıştır. Biyokimyasal ve mikroskobik analizlerin gelişmesiyle mitokondriyal yapı ve mitokondriyal enzim aktiviteleri incelenerek oksidatif ya da glikolitik kapasiteye göre sınıflandırma geliştirilmiştir. Reiser (1985), Staron & Pette (1986) miyozin ağır zincir (MHC) izoformları ile elektroforez, kasılma hızı ve miyozin ATPaz aktivitesi arasındaki ilişkiyi ortaya koymuştur. Schiaffino (1989) ise hızlı iskelet kaslarında bulunan üçüncü lif tipini tanımlamış ve bunu Tip IIX olarak isimlendirmiştir (73). Tablo 2’de lif tipi sınıflandırması özetlenmiştir.
MHC’ler mültigen ailesi tarafından kodlanırlar. Yetişkin memeli kasında 11 farklı MHC izoformu tanımlanmıştır. Bunlardan MHCIβ, MHCIIa, MHCIId ve MHCIIb pek çok kasta yaygın olarak bulunmaktadır. Diğer izoformlar ise sadece özel kas gruplarında
13 bulunmaktadır. Örneğin “süper hızlı” MHCIIm çiğneme kaslarında, MHCeom ekstraoküler kaslarda, MHCIton ekstraoküler ve laringeal kaslarda, MHCIα diyafram, çiğneme kasları ve ekstraoküler kaslarda bulunur (59). Kas lifleri “saf” ya da “hibrid” olabilir. Saf lifler tek tip MHC izoformu içerirken, hibrid lifler iki ya da daha fazla MHC izoformu içerir.
Yetişkin memeli iskelet kasında bulunan başlıca MHC izoformlarına göre kas lifleri; MHCIβ içeren yavaş tip I ve üç hızlı tip; MHCIIa içeren tip IIA, MHC IId içeren tip IID ve MHCIIb içeren tip IIB şeklinde sınıflandırılabilir. MHCIId, MHCIIx’in ve tip IID tip IIX’in eşdeğeridir (44,69). Her iki isimlendirme de literatürde bulunmaktadır. Çalışmalarda memelilerde vücut hacminin arttıkça yavaş MHC izoformlarının ekspresyonunun arttığı gösterilmiştiır. Aslında insan ve diğer büyük memelilerde MHCIIb ekspresyonu yoktur. Önceden insan kasında IIB olarak sınıflanan kas tipi tip IIX ya da tip IID olarak yeniden isimlendirimiştir (21). Hİbrid lifler üç ya da daha fazla MHC izoformu içerebilir. Genellikle dominant olan MHC göre isimlendirilirler. “Yakın komşu” kuralına göre yavaştan hızlıya sınıflanabilirler, saf lifler arasında köprü gibidirler ( tip I - tip I/tipIIA - tipIIA/I - tip IIA - tip IIA/D - tip IID/A - tipIID - tip IID/B - tip IIB/D tip IIB) (59).
Kas lifleri dinamik yapıdadır. Nöromüsküler aktivite, mekanik yüklenmenin artması ya da azalması, hormonal değişiklikler(özellikle tiroid hormonları), yaşlanma gibi nedenlerle fenotipik değişikliklere uğrayabilirler. Kasların innervasyonu değiştiğinde yavaş kasılan kaslar hızlıya, hızlı kasılan kaslar ise yavaşa dönüşürler. Kronik düşük frekanslı uyarı (KDFU) yavaş kasılan kasların uyarımına, fazik yüksek frekaslı uyarı (FYFU) ise hızlı kasılan kasların uyarımına benzer. Hızlı kasılan kaslara KDFU verildiğinde MHC izoformlarının değiştiği ve kasın yavaş tipe dönüştüğü, ratlarda yavaş kasılan soleus kasına FYFU verildiğinde ise hızlı tipe dönüştüğü gösterilmiştir. İnnervasyon değiştiğinde yalnızca MHC değil, troponin subünitleri, tropomiyozin, α-aktinin gibi miyofibriler proteinlerin ve sarkoplazmik retikulum proteinlerinin sentezinde, sitozol Ca düzenleyici sistemde, enzim aktivitesinde, membran proteinlerinde, enerji metabolizmasında da değişiklikler meydana gelmektedir.
Antrenmana uyum ise artan nöromüsküler aktiviteye bağlı gelişir. KDFU’nın etkisine benzer ancak daha küçük bir etki yaratır. Dayanıklılık egzersizinin temel etkisi aerobik-oksidatif enzim aktivitesinin artmasıyla meydana gelir. Germe ve mekanik yüklenmede KDFU gibi hızlı liften yavaş life dönüşe neden olur. Hızlı kaslar gergin pozisyonda immobilize edildiğinde hızlı tip MHC izoformlarının yavaş tipe dönüştüğü, yine hızlı kasılan kasın sinerjik kası cerrahi olarak kesildiğinde meydana gelen yüklenme sonuçunda kasın yavaş lif özelliği kazandığı gösterilmiştir. Yavaş kaslara ise yüklenme azaldığında tip I liflerin sayı ve boyutunun azaldığı ve kasların hızlı kasılan kas özelliği kazandığı, MHC I izoformun azaldığı MHC IIa ve MHC IId izoformlarının arttığı saptanmıştır. Hormonlarda lif fenotipini
14 etkilemektedir. Hipertiroidide kaslar hızlı kasılan tipe kayarken, hipotiroidide yavaş kasılan tipe kayar. Yaşlanma ile hızlıdan yavaşa bir dönüşüm olur. Yaşlanma süresince pek çok içsel ve dışsal faktör bu dönüşümün nedenidir. Merkezi sinir sisteminde gerilemeler (özellikle hızlı α-motor nöron kaybı/periferik sinir sistemi dejenerasyonu), inaktivite, azalan tiroid hormonu seviyeleri bu dönüşüme neden olarak sayılabilir. İnaktivitenin aksonal transportu yavaşlattığı, egzersizin ise aksonal transportu hızlandırdığı gösterilmiştir. Aktivitenin yaşlanma süresince kas-iskelet sisteminin korunmasındaki rolü büyüktür (60).
Lif tipleri arasındaki temel farkı yaratan yukarıda bahsedilen kontraktil proteinlerdir. Diğer önemli iki fark ise Ca döngüsü, sarkoplazmik retikulum (SR) ve mitokondridir. SR kas hücresi volumünün %2-6’sını oluşturur. Bu oran hızlı tip II liflerde en yüksektir (%5-6). Mitokondri yoğunluğu ise yavaş tip I liflerde %6 iken, hızlı tip II liflerde sadece %2-3’tür. Dayanıklılık egzersizi ile üç kas lifi tipinde de mitokondri miktarı artar. Bu artış 4-6 haftalık bir egzersiz ile %30-100’ kadar değişen geniş bir aralıkta meydana gelmektedir (24). Tablo 2’de lif tipi arasındaki temel farklar özetlenmiştir.
15
2.2.2 Enerji Sistemleri
Egzersiz sırasında kas kasılması için gerekli enerji sistemleri üçe ayrılır: ATP-PCr (fosfojen) sistem, anaerobik glikoliz (laktik asit) sistemi ve aerobik sistem.
a. ATP-PCr (Fosfojen) Sistem
Kısa süreli yoğun egzersiz sırasında depo ATP ile birlikte hemen devreye girer. Kas dokusunda kg başına 4-6 mmol ATP ve 15-17 mmol fosfokreatin (PCr) olduğu gösterilmiştir. Ağırlık kaldırma, sprint, tenis servisi gibi saniyelik aktivitelerde depo ATP yeterli olurken, birkaç saniye sürecek maksimal aktivitelerde gerekli ATP’nin yenilenmesi fosfokreatinden sağlanır. Tüm sportif aktivitelerde yüksek enerjili fosfatlar kullanılmasına rağmen, bazılarında sportif performans sadece bu enerji sistemine dayanır. Bu bakımdan kas içi yüksek enerjili fosfat düzeyi kısa süreli aktivitelerde performansı önemli derecede etkiler (26).
b. Anaerobik Glikoliz (Laktik Asit) Sistemi
Kısa süreli yoğun egzersizin devamı için yüksek enerjili fosfatın (ATP) yeniden sentezlenmesi gerekir. Adenozin difosfatın (ADP) fosforilize edilmesi, kas dokusundaki glikojenin pirüvik asitten laktik asite kadar yıkılmasını sağlayan anaerobik (oksijenden bağımsız) glikoliz yolu ile yapılır. Burada 1 mol (180 g) glikojenden 3 mol ATP elde edilir. Antrenman esnasında anaerobik glikoliz ile elde edilen kullanılabilir ATP miktarı aslında daha azdır. Çünkü yorucu bir antrenman esnasında yorgunluk baş göstermeden önce kaslarda ve kanda en fazla 60-70 g laktik asit tolere edilebilir. Eğer 180 g glikojenin tümü tepkimeye girerse 180 g laktik asit oluşur. Bu nedenle kas ve kandaki laktik asit miktarı yorgunluğa eşlik eden düzeye ulaştığında anaerobik glikoliz yoluyla sadece 1-1.2 mol ATP yenilenmiş olur. Anaerobik glikolizin enerji transfer hızı yüksek enerjili fosfat sisteminin %45’i kadardır. Anaerobik glikoliz 1-3 dk süren 400-800 m gibi yarışlarda ağırlıklı olarak devreye girer (26). Bilanço şu şekildedir:
C6H12O6 (GLİKOJEN) → 2 C3H603 (LAKTİK ASİT) + ENERJİ ENERJİ + 3 Pi + 3 ADP→ 3 ATP
c. Aerobik Enerji Sistemi
Bu enerji sisteminde glikoliz ve Krebs döngüsünde ortaya çıkan elektronlar, elektron transfer sistemiyle oksijene iletilir. Bilanço şu şekildedir:
C6H12O6 + 6 O2 → 6 CO2 + 6 H2O + ENERJİ ENERJİ + 39 ADP + 39 Pi → 39 ATP
16 Aerobik metabolizmayla ATP tekrar sentezi için pirüvik asitin direkt olarak Krebs döngüsüne girmesi, yağların β-oksidasyonun ve mitokondri oksijen transferi sistemlerinin devreye girmesi gerekir. Egzersizin süresi 1-3 dakikanın üzerine çıktığında ya da dakikalarca, saatlerce devam ettiğinde genel olarak transfer edilen enerji sistemi aerobik enerji sistemidir (67). Kaslardaki tüm glikojen aerobik yolla yıkılırsa elde edilen ATP miktarı 87-98 moldür. Bu iki anaerobik sistemden elde edilebilecek toplam miktarın yaklaşık 50 katıdır. Ayrıca 80-100 mol glikojen de karaciğerde depolanır. Eğer bunun tümü aerobik metabolizma ile kullanılır ise 17-22 mol daha ATP meydana gelir. Bir sportif aktivitede bu enerji sistemleri, açılıp-kapanma gibi ayrı ayrı değil, aktivite özelliğine (özellikle şiddet ve süre olarak) göre öne çıkarak devreye girmektedir (26).
Şekil 4: Egzersizde enerji kaynakları (24) 2.2.3 Anaerobik güç
Maksimal ve supramaksimal fiziksel aktivite sırasında iskelet kaslarının anaerobik enerji transfer sistemlerini kullanarak meydana getirdiği iş kapasitesi “anaerobik kapasite” olarak tanımlanmaktadır. Güç, birim zamanda meydana getirilen iştir. Anaerobik güç ise birim zamanda anaerobik yoldan yani ATP-PC enerji kaynakları kullanılarak meydana getirilen iştir. Anaerobik iş, patlayıcı gücün ortaya konması anlamına gelen, anaerobik eşik değer üzerinde bir iş yükü olup, erken yorgunluk ile kendini gösteren fiziksel aktivite tipidir. Anaerobik aktiviteye uzun süre devam edilemez. Kas ve kan laktat seviyesi yükselir, biriken laktatın tamponlanması akciğerlerden CO₂ atılımını arttırır ve pH düşmesi (pH=6.4) nedeniyle kaslarda yorgunluk meydana gelir (67).
17 Uzun atlama, yüksek atlama, gülle atma, cirit atma, sürat çıkışları (futbolda, voleybolda, basketbolda) içeren spor dallarında anaerobik güç sporcunun performansını belirler (3). Anaerobik güç, kas içinde ATP yenilenme hızına, kas içi glikojen konsantrasyonuna, yüksek laktik asit ve düşük pH düzeyine toleransa, kas lifi tipi dağılımına ve kas koordinasyonu gibi özelliklere bağlı olarak ortaya konmaktadır. Bu nedenle anaerobik gücü direkt olarak ve objektif ölçen tek bir test yoktur. Ölçüm, anaerobik gücü kısmen yansıtacak testler ile yapılabilir (22). Anaerobik güç testleri; Margaria-Kalamen basamak testi, Wingate testi, Durarak dikey (Sargeant) sıçrama testi, Bosco sıçrama testi, 50 yarda (45 m) koşu testi olarak sayılabilir. Futbolda üst düzey performans için sıçramanın anahtar rol oynadığı bilinmektedir ve sıçrama mesafesi ile sürat performansı arasında yüksek korrelasyon olduğu gösterilmiştir (82). Futbolcular ile yapılan önceki çalışmalarda eller serbest dikey sıçrama 47.8-60.1 cm, eller belde dikey sıçrama 38.8-42.2 cm, squat sıçrama 37.0-40.2 cm, durarak uzun atlama 225 cm olarak ölçülmüştür (4,50,75,77).
2.2.4 Aerobik Güç / Maksimal Oksijen Kullanımı
Kas dokusuna taşınan oksijeni bu dokunun maksimal kullanım kapasitesidir. Bu nedenle aerobik güç akciğerler, kardiyovasküler sistem ve hematolojik bileşenlerinin fizyolojik kapasitelerine ve egzersiz sırasında aktif olan kasların oksidatif mekanizmalarının etkinliğine bağlıdır. Kişinin birim zamanda kullanabildiği oksijen miktarı ne kadar yüksek ise aerobik gücü de o kadar yüksek demektir. Aerobik güç dayanıklılık sporlarında performansa etkili en önemli fizyolojik faktördür. Maksimal oksijen kullanımından ilk bahseden A.V. Hill olmuştur. Kişiye giderek artan bir iş yaptırıldığında kullanılan oksijen miktarı lineer bir şekilde artar ve sonunda öyle bir noktaya gelinir ki, yapılan iş artsa dahi oksijen kullanımı aynı düzeyde kalır. Bu oksijen düzeyi Maks VO₂ (maksimal oksijen kullanımı) olarak adlandırılır. Kişinin bu değeri dakikada litre veya ml cinsinden total miktar olarak ifade edilebilir ancak fizyolojik karşılaştırılabilir bir ifade olması için vücut ağırlığının kilogramı başına düşen miktarı hesaplamak ve ml/dk/kg ya da L/dk/kg olarak ifade etmek daha doğrudur (3,22).
Maks VO₂ aşağıdaki şekilde formüle edilebilir:
Maks VO₂ = Kalp debisi (Kalp atım hızı x atım hacmi) x a-v O2 farkı
Eşitliğin sağ tarafındaki üç faktördeki değişiklik Maks VO₂’de de değişikliğe neden olur. Bu nedenle değişkenlerin ayrıntılarından bahsetmek gereklidir.
a. Kalp Debisi
Kalp debisi, bir dakikada perifere gönderilen kan miktarı olarak tanımlanmaktadır. Atım hacmiyle nabzın ilişkisine dayalı bir şekilde hesaplanabilmektedir. Atım hacmi (stroke volume), kalbin bir kasılma sırasında perifere gönderdiği kan miktarını ifade etmektedir ve
18 yaklaşık 70-90 ml kadardır. Sağlıklı bir kişinin sinoatrial düğümünden bir dakikada çıkan uyarı sayısı ortalama 75 (60-90), kalp debisi ise dinlenik durumda yaklaşık beş litre kadardır.
Kalp debisi (Q) = Atım hacmi x Kalp atım hızı
b. Atım Hacmi
İstirahatte 70-90 ml iken, maksimal bir egzersizde 120-155 ml’ye kadar çıkabilir. Her sistolde kalbin pompaladığı kan miktarı iki faktöre bağlıdır:
1. Sistol başında ventrikülün içerdiği kan volümü: buna diyastol sonu volüm adı verilir. 2. Ventrikülün kasılarak boşalabilme derecesi: buna da sistol sonu volüm denir. Diyastol sonu volüm ile sistol sonu volüm arasındaki fark atım volümünü verir. Egzersiz esnasında kalbin atım volümünün artması ventriküllere venöz dolaşımdan daha fazla kan gelmesi yani diyastol sonu volümün artmasına (Starling Yasası) ve ventriküllerin daha kuvvetli kasılarak sistol sonu volümün daha fazla küçülmesine bağlıdır. Diyastol sonu volüm venöz dönüşe bağlıdır. Venöz dönüşü etkileyen faktörler şunlardır:
Venlerin refleks vazokonstriksiyonu: egzersiz sırasında periferik venlerin tonusu artar, refleks vazokonstriksiyon meydana gelir böylece kan periferde göllenmez, venöz dönüş kolaylaşır.
Kasların pompa etkisi: kaslar kasıldığında içlerinden geçen venleri sıkıştırılar ve kan kalbe doğru pompalanır, venlerdeki tek tönlü kapaklar sayesinde kas gevşediğinde kanın geri dönüşü engellenir. Koşu gibi ritmik egzersizler sırasında bu mekanizma oldukça etkili çalışırken, izometrik ve statik egzersizler venöz kanın kalbe dönüşünü engeller.
Solunum pompası: venöz dönüş inspirasyonda intratorasik basınç azaldığı için artar, ekspirasyonda ise azalır.
Atım hacminde diyastol sonu volümün etkisi önemlidir, ancak ventrikülün sistol sonu volümü yani ventrikülün kasılma kuvveti daha önemlidir. Egzersiz esnasında artan epinefrin ve sempatik aktivite miyokardın kasılma gücünü arttırır. Atım hacmi istirahatte sedanter bireylerde 70 ml civarındayken, egzersiz sırasında 120-125 ml’ye yükselir, antrene bireylerde ise 150 ml, hatta aerobik kapasitesi çok yükselmiş sporcularda 200-210 ml’ye kadar çıkabilir (3).
c. Kalp Atım Hızı
Sempatik-parasempatik sinir sisteminin kontrolünde olan kalp hızı, dolaşım fonksiyonunun izlenmesinde önemli bir parametredir. Antrenmanlı bireylerde atım hacmi arttığı için istirahat nabzı düşer, yine belirli iş yükünde sedanterlere göre nabız daha düşüktür (22).
19 İstirahat Q = Atım hacmi x Kalp Atım Hızı
Antrenmansız Q = 4.9 L = 70 ml x 70 vuru/dk Antrenmanlı Q = 4.9 L = 100ml x 49 vuru/dk
Her bireyin maksimal nabız değeri vardır ve genellikle “220-yaş” formülü ile hesaplanır. Bu formül yaklaşık tahmini maksimal kalp hızını verir ve ± 10 standart sapması vardır. Efordan sonra nabzın normale dönmesini iki faktör belirler: egzersiz sırasındaki iş yükü ve kişinin kondisyon düzeyi. Antrenmanlı bireylerde nabız daha kısa sürede normale döner (3).
Tablo 3: Antrenmanlı ve antrenmansız bireylerde oksijen taşıma sisteminin bileşenleri (26)
Koşullar VO₂ (ml/dk) Atım hacmi (L/vuru) Atım sayısı (/dk) a-v O2 farkı (ml/L) İstirahat Antrenmansız 300 0.075 82 48.8 Maksimal Egzersiz Antrenmansız 3100 0.112 195 138 İstirahat Antrenmanlı 300 1.105 58 49.3 Maksimal Egzersiz Antrenmanlı 3440 0.126 192 140.5 Maksimal Egzersiz Maratoncu 4470 0.159 190 155
d. Arterio-Venöz Oksijen (a-v O₂) Farkı
Arter kan oksijen miktarından venöz kan oksijen miktarı çıkarıldığında elde edilen değer arterio-venöz oksijen farkı olarak isimlendirilir. İstirahatte kasa gelen 100 ml arteriyel kanda oksijen miktarı 20 ml iken, kası terk eden oksijen 15 ml’dir, yani istirahatte arterio-venöz oksijen farkı yaklaşık 5 ml’dir. Egzersiz sırasında kan pH’sındaki düşüş, kan karbondioksit konsantrasyonundaki artış, 2-3 difosfogliserat konsantrasyonunu artışı, vücut ısısındaki artış gibi faktörler kasa oksijen bırakılmasını kolaylaştırır. Şiddetli egzersizler sırasında arterio-venöz oksijen farkı 15-17 ml’ye kadar çıkabilir (3). Arterio-venöz oksijen farkını etkileyen faktörler hemoglobin miktarı, kapiller yoğunluk ve mitokondri miktarıdır. Kapiller sayısının artması, kapiller-mitokondri difüzyon mesafesini kısaltır ve kanın kapiller geçiş süresini uzatır. Böylece kandan daha fazla oksijen çekilmesi sağlanır, bu oksijenin kullanılması için mitokondri sayısının da artması gerekmektedir.
20
Şekil 5: Antrenmanlı ve antrenmansız bireylerde egzersizin a-v O₂ farkına etkisi: Antrenmanla kas kandan daha fazla oksijen alır ve kullanır (3)
Egzersiz esnasında kalbin bir dakikada tüm organizmaya gönderdiği kan miktarı artar ve bunun dokulara dağılımı, dokuların gereksinimine fizyolojik bir uyum gösterir. İstirahat halinde iskelet kaslarına giden kan, kalbin dakika volümünün %15-20’ si iken, egzersizde bu oran %85-88’e yükselir. Abdomene giden kan azalır, beyine giden kan miktarı değişmez.
2.2.5. Maksimal Oksijen Kullanımına Etki Eden Faktörler
Cinsiyet
Maks VO₂ değeri ile yağsız vücut kitlesi arasında anlamlı korrelasyon olduğu bilinmektedir. Bu nedenle Maks VO₂, erkeklerde kadınlardan %20-25 daha yüksektir. Erişkin bir kadında vücut yağ oranı %26, erkekte %15 civarındadır. Maks VO₂ yağsız vücut kitlesine göre hesaplandığında erkek ve kadın arasındaki aerobik kapasite farkının oldukça düşük olduğu görülür. Bu küçük farkın nedeni kadınlarda hemoglobinin daha düşük olmasıdır. Kadın maratoncuların incelendiği bir çalışmada kadınların Maks VO₂ değeri erkeklere göre %16 düşük çıkmış, ancak yağsız vücut kitlesine göre kıyaslandığında bu fark %7.8’e düşmüştür (81).
Yaş
Doğumdan itibaren Maks VO₂ artar. Astrand’a göre Maks VO₂ 18-20 yaşında zirveye ulaşır, sonra giderek azalmaya başlar. Sedanterlerde 25 yaş sonrası, Maks VO₂ değeri her yıl %1 azalmaktadır. Katılımcıların yaş ortalamasının 70 olduğu altı yıl takipli bir çalışmada aerobik gücün erkekte %24/dekat, kadında %18/dekat düştüğü gösterilmiştir (37). Benzer bir
21 çalışmada gençlerde (20-30 yaş) Maks VO₂ kaybı %5/dekat, yaşlılarda (≤60yaş) ise %20/dekat bulunmuştur (25). Yaşlanmayla birlikte kalp atım hızı her dekatta %3-5 azalır, atım hacmi düşer; a-v O₂ farkının da her dekatta %3 azaldığı gösterilmiştir. Ayrıca yaşlandıkça aerobik kapasitenin azalmasına neden olan faktörler; kas kütlesi azalması, yağ dokusu artması, periferik rezistansın artması, kas kapiller yoğunluğunun azalması ve endotelyal disfonksiyon olarak sayılabilir (28). Yaşlı kişilerde sedanter yaşam şeklinin getirdiği değişiklikler de bu azalmada etkilidir, düzenli egzersiz ile bu azalma yavaşlatılabilir.
Kondisyon Düzeyi
Düzenli yapılan aerobik egzersizlerin seviyesi, Maks VO₂ üzerinde önemli etkiler yaratır. Bir çalışmada altı ay boyunca Maks VO₂’nin %75 ile haftada 3-4 gün jogging yapıldığında Maks VO₂’nin 40.5 ml/dk/kg’dan 49.8 ml/dk/kg’a çıktığı gösterilmiştir. Tablo 4’de çeşitli spor branşlarındaki Maks VO₂ değerleri verilmektedir.
Genetik
Maks VO₂’yi etkileyen en önemli faktörlerden biri de kalıtımdır. Bouchard, 86 çekirdek aile üzerinde yaptığı araştırmada genetiğin Maks VO₂’ye etkisinin yaklaşık %50 olduğunu göstermiştir (10). Çeşitli çalışmalarda; 10 monzigot ikizin 20 hafta, 8 ikizin 3 ay ve 6 ikizin 15 hafta antrenmanı öncesi ve sonrasında aerobik gücü ölçülmüş ve Maks VO₂ düzeyindeki artışın ikiz eşleri arasında oldukça benzer olduğu gösterilmiştir (12,32,63).
22
2.2.6 Anaerobik Eşik
Bireyin fiziksel iş kapasitesinin değerlendirilmesinde Maks VO₂ önemli bir kriterdir, ancak tek belirteç değildir. Uzun süre egzersiz yapabilme kapasitesi Maks VO₂’den çok, belirli bir egzersiz şiddetindeki kan laktik asit konsantrasyonu ile daha yakın ilişkilidir (22). Anaerobik eşik kanda laktik asit birikmesinin hızlandığı, efor için gerekli enerjide anaerobik süreçlerin payının arttığı efor düzeyidir. Egzersiz yoğunluğu yükseldiğinde, ATP yenilenmesi anaerobik metabolizmayla desteklenir. Anaerobik eşik Maks VO₂’nin %60’ı kadardır. Dayanıklılık antrenmanlarının tek etkisi Maks VO₂’yi arttırmak değil, aynı zamanda organizmayı Maks VO₂’nin büyük kısmını daha az laktik asit birikimi ile kullanılır duruma getirmesidir. Dayanıklılık sporcularında bu oran Maks VO₂’nin %85-90’na kadar çıkabilir (3).
İş yükü giderek arttırılarak yapılan egzersiz testinde anaerobik eşik değer noninvazif gaz değişim yöntemleriyle saptanırsa, “Anaerobik eşik değer” veya “Metabolik eşik değer” olarak tanımlanır (V-Slope yöntemi). Laktat değerleri saptanarak ölçülürse “Laktat birikim eşik değer” tanımı kullanılır. Anaerobik eşik değer ölçümü, oksijen alımı ile doğrusal olmayan pulmoner ventilasyondaki artış prensibine dayanır. Şiddeti artan egzersiz testlerinde, egzersizin başlangıcında VCO₂, kasların kullandığı oksijen miktarına yanıt olarak oluşur. Egzersiz süresi ilerleyince, kasların yükü artar ve VCO₂, oksijen kullanımına yanıt olarak değil, kan laktatının tamponlanması sonucu yükselmeye başlar. VCO₂ gittikçe VO₂ eğrisinden uzaklaşır. Bu iki eğrisinin kırılma noktasına “Anaerobik eşik değer” denir. Bu noktada laktat birikmeye başlar. Laktat düzeyi iş yoğunluğuna paralel olarak yükselmeye devam eder. Solunumsal kompansatuar mekanizma nedeniyle hiperventilasyon meydana gelir. Yoğun egzersizde sempatik aktivitenin artması, karaciğer, böbrek gibi organlardaki iskemi kan laktat oluşumunun daha da artmasına neden olan faktörlerdir.
Kan laktat eşik değerinin 4.0 mmol/l olarak kabul edilmesine karşılık, çalışmalarda kan laktat eşik değerinde oldukça geniş ölçüde kişisel farklılıklar bulunduğu gösterilmiştir (2.1-12.6 mmol/l). Anaerobik eşik değer aerobik kapasite derecesinin önemli göstergesidir. Sporcularda antrenman programlarının düzenlenmesinde antrenman yoğunluğunun belirlenmesinde önemlidir (67).
23
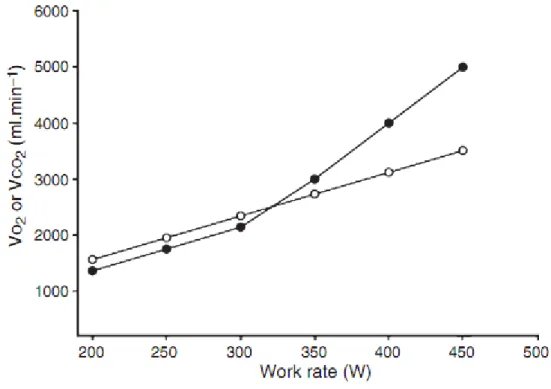
Şekil 6:Artan iş yükünde tüketilen oksijen hacmi(VO₂) ve üretilen karbondioksit hacmi (VCO₂), VO₂(○)’de lineer artış VCO₂(●)’de bifazik artış (39).
2.2.7 Maksimal Oksijen Kullanım Kapasitesi Ölçümü
Maks VO₂ direkt ya da endirekt yöntemler ile ölçülebilir. Endirekt ölçüm yöntemlerinde Maks VO₂, submaksimal yüklemeyle kalp hızı, yük, zaman, mesafe vb. parametre değişiminden hesaplanır. Bu nedenle maksimal ölçümün riskli olduğu kişilerde daha güvenlidir ve yöntem olarak uygulamak daha kolaydır. Ancak Maks VO₂ saptamanın en doğru yolu direkt ölçüm yöntemidir. Direkt ölçüm; laboratuvar koşullarında maksimal yüklenmede, solunum havasındaki O₂ ve CO₂’in gaz analizörleriyle analizi prensibine dayanır. Maks VO₂ ölçümü, ergometre üzerinde (bisiklet, koşu bandı gibi) düşük iş düzeyinde başlar. Isınma için bir zaman verilir, önceden belirlenmiş test protokolleri kullanılarak yoğunluğu giderek artan yüklenme uygulanır ve ekspire edilen gazların analizi yapılır. Kişi, giderek artan iş yaptığında kullandığı O₂ miktarı doğrusal bir şekilde artar. Bir ünite zamanda alınan O₂ dolaşım ve solunum sistemlerinin sınırlaması nedeniyle belirli bir maksimum düzeye ulaşır ve bu noktadan sonra yapılan iş artsa da O₂ kullanımı artık daha fazla bir artış göstermez, aynı düzeyde kalır ya da düşme eğilimi gösterir. Ekspire edilen hava test boyunca toplanır, tüketilen oksijen miktarı ölçülür ve tüketilen en yüksek değer Maks VO₂ olarak adlandırılır.
24
2.2.8 Futbolda Dayanıklılık ve Sürat Performansı
Futbol; çeviklik, esneklik, elastikiyet, denge, kassal ve kardiyorespiratuar dayanıklılık, koordinasyon gibi faktörlerin performansa beraberce etki ettiği bir spor disiplinidir. Üst düzey bir futbolcunun oyun esnasında yaklaşık 9-14 km mesafe kat ettiği gösterilmiş, maç boyunca 1330 yer değiştirmenin 220’si yüksek hızda gerçekleşmiştir. Barros ve ark. Brezilya liginde yaptıkları bir çalışmada maç boyu toplam kat edilen mesafeyi ortalama 10 km bulmuşlar, yürüme ve jogging tempolu koşu 5.5 km, yavaş tempolu koşu 1.6 km, orta tempolu koşu 1.7 km, yüksek hızlı koşu 0.7 km ve sprint ise 0.4 km bulunmuştur (6,67). Bu sonuçlara bakıldığında futbolda performansı hem anaerobik, hem de aerobik güç belirlemektedir denebilir. Futbolcularda ortalama Maks VO₂ değeri 55-67 ml/dk/kg bulunmuştur, ancak 70 ml/dk/kg üzerinde ölçülen değerler de vardır (36). Bu değerler uzun mesafe koşucularından düşük, sprinterlerden yüksektir. Ayrıca; İngiltere birinci liginde dikey sıçrama 56.0 cm, durarak uzun atlama 2.20 m bulunmuştur. İskoç futbolunda üst düzey sporcularda dikey sıçranma 58.4 cm, durarak uzun atlama 2.44 m olarak saptanmıştır (40).
2.3 Telomeraz Enzimi
Telomeraz enziminin yapı ve fonksiyonunu anlayabilmek için öncelikle telomerden bahsetmek gereklidir.
2.3.1 Telomer
Telomerler, ökaryotik organizmalarda kromozomların uçlarında bulunan, kodlanmayan özelleşmiş DNA tekrar dizilerinden (memelilerde TTAGGG) oluşan heterokromatik bölgelerdir. Kromozomların ucunda yalıtımı sağlamak için yer alırlar. Yani insanda 46 kromozom olduğuna göre ve her kromozom her iki ucunda telomer taşıdığına göre; bir insan DNA’sında toplam 92 adet telomer vardır (1). DNA replikasyonu sonucunda bütün kromozomların sonlarında bir eksilme olduğu ve belli bir sayıda bölünme yapabildiği ve kritik bir eksilme noktasından sonra hücrenin ölümüne neden olduğu gösterilmiştir (5). Yaşlanmayı inceleyen Leonard Hayflick insan fibroblastlarının kabaca 50 hücre bölünmesi sonunda gelişme ve bölünme yeteneklerini yitirdiğini bulmuştur (34). Normal memeli somatik hücreleri, in vitro koşullarda belli sayıda bölünebilir. Bu maksimum bölünme sayısına “Hayflick Limiti” ya da “Proliferasyon Limiti” denir.
Telomerlerin fonksiyonları; replikasyonda lineer kromozomal DNA molkülünün son kısmının tammalanmasında rol oynar. Kromozomların bütünlük ve kararlılığını sağlar; kromozomu yıkım, rekombinasyon ve füzyon gibi durumlara karşı korur, kromozomların nükleus zarına tutunarak belirli bir pozisyonu korumasını sağlar (5).
25
2.3.1.1 Telomerin Yapısı
Telomerin DNA’sı CA-GT açısından zengin basit tandem tekrarlardan oluşur. Telomer subtelomerik ve esas telomerik bölge olaraka ikiye ayrılabilir. Subtelomerik bölgenin boyu değişkendir ve DNA tekrarları heterojendir. İnsan kromozomunda subtelomerik bölge 40-60 kilobaz kadar olup TTGGGG, TGAGGG gibi farklı tekrarlardan oluşur. Esas telomerik bölge ise 5-15 kilobaz kadar olup sadece TTAGGG tekrarından oluşur. Telomerler büyük terminal “loop”lar şeklinde sonlanır.
Şekil 7: T-loop ve D-loop yapısı (52)
Telomer ucundaki TTAGGG dizisi D-loop içinde yer alan CCCTAA dizisi ile baz eşleşmesi yapar ve T-loop oluşturulur. Böylece oluşan T-loop DNA’nın ucunu kırıklardan korur. Ayrıca telomerlerin yapısında telomere bağlı proteinler yer alır. Bu proteinler telomer ucunu sarar, T-loop oluşumuna yardım eder ve telomeri yıkılmaktan korur. Ayrıca telomeraz enziminin telomere bağlanıp bağlanmamasını sağlayarak telomer boy uzunluğunu kontrol eder (52).
2.3.2 Telomeraz
Telomeraz, ökaryotik kromozomların telomerlerine telomerik tekrarların eklenmesini katalizleyen ribonükleoprotein yapıda bir revers transkriptazdır ve büyük bir enzim kompleksidir (1). Telomerlerin korunması için telomeraz aktivitesine gereksinim vardır, çünkü DNA polimerazlar düz DNA uçlarını tam olarak replike edemezler. Telomeraz enzimi, Greider ve Bluckburn tarafından 1985 yılında keşfedilmiştir. İnsanda ise telomeraz aktivitesine servikal kanser hücre hattında rastlanmıştır. Bunun dışında fetal, yeni doğan ve yetişkin testis ve overlerinde rastlanmış, fakat yaşlıların testislerinde bulunmamıştır. İnsanın fetal dokularında, yeni doğanların periferal kan hücrelerinde yüksek oranda telomeraz aktivitesi görülür. Yetişkinlerde, tek çekirdekli periferal kan hücrelerindeki telomeraz aktivitesi tümör hücreleri ve çocukların telomeraz aktivitesine göre daha düşüktür. Özellikle 19 yaşından daha yaşlı bireylerde telomeraz aktivitesi düşmektedir. Telomeraz aktivitesi, bir çok insan
26 somatik dokusunda görülmez. Genellikle yüksek replikatif kapasitesi olan dokularda ve bir çok insan kanser türünde görülür (52).
2.3.2.1 Telomerazın Yapısı
Telomeraz, RNA ve proteinler olmak üzere iki kısımdan oluşur. Telomeraz aktivitesi, yapısında yer alan RNA alt birimine bağlıdır. Bu RNA’ların hepsi protein kısmının avuç içi kısmında uzanan ve yaklaşık olarak 1.5 telomer tekrarına komplemanter olan bir diziyi içermektedir. Bu dizi insanda 5’UAACCUAA3’ şeklindedir.
Şekil 8: Telomeraz bileşenleri (52)
Telomeraz proteinleri ise telomeraz revers transkriptaz (TERT), TEP1 (telomerase associated protein1), p23, hsp90, dyskerin, L22, La autoantigen, hStau, hnRNPs olarak sayılabilir. TERT farklı canlılarda farklı isimler almaktadır, insanda hTERT adını alır. Yapısı yarı açık sağ ele benzetilmektedir. Enzim kompleksinin taşıdığı RNA’nın avuç içi kısmında yer alan 1.5 telomerik tekrara tamamlayıcı olan kısmı DNA’nın G3 ucunun uzamasında görev alırken; RNA’nın geri kalan kısımlarının görevi tam olarak bilinmemektedir. İn vitro çalışmalarda telomeraz aktivitesi bulunmayan insan hücrelerinde, HTERT proteininin sentezlenmesi, telomerazı tekrar aktif hale getirmiştir (31).
Şekil 9: Yarı açık el yapısı (52)
Telomeraz enzimi bütün bu yapıları kullanarak telomerin uzamasını sağlar. Bir çeşit revers transkriptaz olan telomeraz enzimi yapısında bulunan RNA’sını kalıp olarak kullanarak telomerleri uzatır.