Scientific paper
The ultrastructural research of liver in experimental obstructive
jaundice and effect of honey
Bulent Kılıcoglu, M.D.
a,*, Cem Gencay, M.D.
a, Kemal Kısmet, M.D.
a,
Sibel Serin Kılıcoglu, M.D.
b, Imge Erguder, M.D.
c, Serap Erel, M.D.
a,
Asli Elif Sunay, Ph.D.
d, Esra Erdemli, M.D.
e, Ilker Durak, Ph.D.
c, Mehmet Ali Akkus, M.D.
a aAnkara Training and Research Hospital, 4th General Surgery Department, Ankara, TurkeybUfuk University School of Medicine, Department of Histology and Embriology, Ankara, Turkey cAnkara University School of Medicine, Department of Biochemistry, Ankara, Turkey
dFood Engineer, Balparmak Pazarlama, Cekmekoy, Istanbul, Turkey
eAnkara University School of Medicine, Department of Histology and Embriology, Ankara, Turkey
Manuscript received Febraury 7, 2007; revised manuscript April 10, 2007
Abstract
Background: To examine the effects of honey on oxidative stress and apoptosis in experimental obstruc-tive jaundice model.
Method: Thirty rats were divided into 3 groups: group I, sham-operated; group II, ligation and division of the common bile duct (BDL); group III, BDL followed by oral supplementation of honey 10 g/kg/d. Liver samples were examined under light microscope and transmission electron microscope. Hepatocyte apoptosis was quantitated using the terminal deoxy-nucleotidyl transferase-mediated deoxyuridine triphos-phate nick-end labeling (TUNEL) assay. Plasma and blood malondialdehyde (MDA) and glutation activities were measured for determining the oxidative stress.
Results: The liver levels of MDA and GSH were significantly different between the honey and BDL groups (P ⫽ .006 and .001, respectively). However, there was no significant difference between the plasma MDA and GSH levels of these groups (P ⬎ .05). In group III, significant reductions in the size of enlarged hepatocytes and the edema were demonstrated. The dilatation of the bile canaliculi dramatically turned to original dimention. By TUNEL assay, it was shown that administration of honey decreased the number of apoptotic cells. Conclusions: In the present study, we found that honey diminished the negative effects of BDL on the hepatic ultrastructure. We conclude that this effect might be due to its antioxidant and anti-inflammatory activities. © 2008 Excerpta Medica Inc. All rights reserved.
Keywords: Honey; Hepatic apoptosis; Obstructive jaundice; Oxidative stress
Apoptosis or programmed cell death is an essential physi-ologic process that plays a critical role in development and tissue homeostasis. However, apoptosis is also involved in a wide range of pathological conditions, including acute neu-rologic injuries, neurodegenerative diseases, cardiovascular diseases, immunologic diseases, acquired immunodefi-ciency syndrome, portal vein occlusion, biliary obstruction, and cancer. Apoptotic cells may be characterized by specific
morphologic and biochemical changes such as cell shrink-age, chromatin condensation, and internucleosomal cleav-age of genomic DNA. At the molecular level, apoptosis is regulated by the activation of the aspartate-specific cysteine protease (caspase) cascade [1].
The liver has the unique ability of eliminating damaged cells by necrosis or apoptosis, and maintaining its volume through regeneration. The atrophy– hypertrophy complex occurs in the liver when bile flow or portal venous blood flow to a portion of the liver is impaired. The pathophysi-ologic response of the liver in this condition is atrophy of the involved hepatic area and hypertrophy of the uninvolved area. Histopathologically, the atrophy– hypertrophy com-plex is characterized by septal fibrosis in the atrophic liver
Supported by Balparmak Pazarlama Koll. Sti., Istanbul, Turkey. * Correspondıng author. S.B. Ankara Egitim ve Arastirma Hastanesi 4, Cerrahi Klinigi 06340, Ulucanlar/Ankara, Turkey. Tel.:⫹90 312 595 34 49; fax:⫹90.312.363 33 96.
E-mail address: [email protected]
0002-9610/08/$ – see front matter © 2008 Excerpta Medica Inc. All rights reserved. doi:10.1016/j.amjsurg.2007.04.011
with biliary piecemeal necrosis, apoptosis, and ductular proliferation. The atrophic region is marked by widening of the sinusoids[2].
Cholestasis results in the accumulation of toxic bile salts in the liver, which induce Fas-mediated apoptosis by oli-gomerization of the Fas death receptor, activation of Fas-associated death proteins, and activation of the caspase cascade. Fas-mediated apoptosis is important soon after biliary obstruction, and redundant apoptotic pathways can be induced in the presence of impaired Fas activation[2].
Oxidative stress, defined as an imbalance between radi-cal generating systems and radiradi-cal scavenging systems giv-ing rise to free radical induced tissue damage, occurs in patients with liver disease. Reactive oxygen species include superoxide anions, hydroxyl radicals, hydrogen peroxide, and hydroxyethyl radicals, and are generated from a variety of insults such as drug and toxin metabolites, ischemia– reperfusion, cholestasis, and alcohol metabolism. Reactive oxygen species are involved in necrosis and apoptosis of hepatocytes, and contribute to hepatic stellate cell activa-tion. Several major classes of free radical scavengers, such as superoxide dismutase, catalase, and glutathione peroxi-dase, were investigated in various forms of liver injury. These substances afforded effective protection against the oxidative insults to the tissue. Bile acids cause oxidative damage by stimulating the generation of oxygen free radi-cals from mitochondria, as well as promoting their release from neutrophils and macrophages [3,4]. Besides chemo-taxis of inflammatory cells, monocyte chemoattractant pro-tein-1 and macrophage inflammatory protein-2 can stimu-late the release of reactive oxygen species from Kupffer cells and neutrophils [5]. Cholestasis augments inflamma-tory responses to gram-negative endotoxemia, sensitizing the host to enhanced fluid flux in multiple organs and to mortality by a leukotriene-independent mechanism. Circu-lating tumor necrosis factor (TNF)-␣ and interleukin (IL)-6 are significantly enhanced in rats with chronic biliary cho-lestasis[6]. Moreover, biliary obstruction in the rats results in a significant depression of reticuloendothelial system phagocytic function[7].
Honey is a supersaturated sugar solution produced by honey bees from nectar of different plants. Fructose and glucose are the main components, with other chemical com-pounds in small quantities, including sucrose, glucose oxi-dase, hydrogen peroxide, phenolics, flavonoids, and ter-penes. It has a long tradition of use for wound healing since ancient times. It was used alone or in combination with other substances externally or orally. Honey has bacteri-cidal, bacteriostatic, antifungal, antiviral, scolibacteri-cidal, antiox-idant, antitumoral, hepatoprotective, and anti-inflammatory effects[8 –22].
We examined the effects of honey on oxidative stress and apoptosis in obstructive jaundice model in this study.
Materials and Methods
Animals
Thirty Wistar-Albino male rats, weighing 250 ⫾ 25 g, were housed under constant temperature (21 ⫾ 2°C) indi-vidually in wire cages under a 12-hour light-dark cycle. Twelve hours before anesthesia, animals were deprived of
food but had free access to water 2 hours before anesthesia. No enteral or parenteral antibiotics were administered at any time. The rats that died during the experiment were ex-cluded from the study and no new rat was inex-cluded. The procedures in this experimental study were performed in accordance with the National Guidelines for The Use and Care of Laboratory Animals and approved by Animal Ethics Committee of Ankara Research and Training Hospital. Study groups
Rats were randomly divided into 3 groups each including 10 animals: group I, sham-operated; group II, ligation and division of the common bile duct (BDL); group III, BDL followed by oral supplementation of honey 10 g/kg/d (Bal-parmak Ltd, Istanbul, Turkey) with a nasogastric tube that was inserted daily and taken off after honey supplementa-tion. Animals were killed on postoperative day 7 by high-dose diethyl ether inhalation. Blood and liver samples were taken for examination.
Operative procedure
Animals were anesthetized by intramuscular injection of 30 mg/kg ketamine hydrochloride (Ketalar; Parke-Davis, Istanbul, Turkey) and 5 mg/kg xylasine (Rompun; Bayer, Istanbul, Turkey). Midline laparotomy was performed under sterile conditions. In the sham-operated group (group I) the common bile duct (CBD) was freed from the surrounding soft tissue and was manipulated without ligation and tran-section. In group II and III, CBDs of the rats were identified, double ligated with 5-0 silk, and divided between the liga-tures. The same surgeon performed all procedures. The abdominal incisions were closed in 2 layers with continuous 3-0 silk sutures. Animals were allowed to feed after the operation.
Evaluation of oxidative stress
Postmortem liver samples were taken and kept on an ice bath until homogenization. The tissues were homogenized in (20 wt/vol) serum physiologic solution, then centrifuged at 4,000g for 15 minutes and upper clear supernatants were used in the assays[23]. All the procedures were performed at 4°C throughout the experiments. Protein level of the clear supernatants was studied by Lowry’s method[24]. Malon-dialdehyde (MDA) levels (nmol/mg) and glutation-peroxi-dase (Px) (mIU/mg) enzyme activities were measured in the supernatants. MDA levels were measured by thiobarbituric acid reactive substances method[25]. After the samples were preincubated with fish oil and xanthine– oxidase system at room temperature for 1 hour, MDA level was determined. GSH-Px activity was measured by following changes in NADPH absorbance at 340 nm[26]. Plasma levels of MDA and GSH were determinated by the same method.
Histopathologic examination
The histopathological analyses were performed in the Histology and Embriology Department of Ankara Univer-sity School of Medicine. For light microscopic analyses, the samples were obtained from the liver, fixed in 10% neutral buffered formaline solution for 2 days. Tissues were washed in flowing water and were dehydrated with rising
concen-trations of ethanol (50%, 75%, 96%, 100%). After dehydra-tion, specimens were put into xylene to obtain transparency and were then infiltrated with and embedded in paraffin. Embedded tissues were cut into 5-m thick sections by Leica RM 2125 RT (Bensheim, Germany) and stained with hematoxylin and eosin, and Mallory-Azan dyes. His-topathologic examinations were performed by 2 his-topathologists blinded to the study design. Photographs were taken by Nikon eclipse E 600 (Japan).
For transmission electron microscope (TEM) analyses, samples were fixed with phosphate buffered (pH 7.3) 2.5% glutaraldehyde and 2% paraformaldehyde mixture solution for 2 hours at room temperature. They were washed with phosphate-buffered saline solution (PBS) (pH 7.3) and were fixed with 1% osmium tetraoxide for 2 hours as secondary fixation. After washing, they were embedded in Araldite 6005 (EMS 10920, Washington) and were cut with Leica EM FCS (Vienna, Austria) ultramicrotome. Semi-thin 1-m sections were stained by toluidine blue–azur II to select region of interest for the following procedures. Sixty- to 70-nm thin sections were stained with uranil acetate and lead citrate. They were examined and photographed using a LEO 906 E TEM (80 kV; Oberkochen, Germany).
TUNEL assay
Hepatocyte apoptosis was quantitated using the terminal deoxy-nucleotidyl transferase-mediated deoxyuridine triphos-phate nick-end labeling (TUNEL) assay. All liver tissue spec-imens were fixed in freshly prepared 3.5% paraformaldehyde and sucrose. Tissues embedded in Thermo Shandon Cryoma-trix (catalog no. 6769006, UK) and sectioned (10m) using a Shandon Cryotome, then saved at 4°C. After treating the specimen with PBS, the sections were processed following the instruction of a commercial kit (Roche D-68298, In situ Cell Death Detection kit fluorescein, catalog no. 1 684 795; Mannheim, Germany). After rinsing the specimens twice with PBS, they were mounted with H-1398 Hoescht, 33258 Molec-ular Probes (USA). The number of TUNEL-positive cells was counted in 10 random microscopic fields (100⫻). Each micro-scopic field contained approximately 970⫾ 35 hepatocytes.
Statistical analysis
Data analysis was performed using the Statistical Pack-age for Social Sciences (SPSS) version 11.5 for Windows (SPSS Inc, Chicago, IL). According to Shapiro Wilks’ test all variables, except for mean number of TUNEL assays,
were normally distributed about the mean. Data were pre-sented as means⫾ SD. Differences among the groups were evaluated by one-Way analysis of variance (ANOVA) or Kruskal-Wallis variance analysis, whichever was appropri-ate. When the P values from the variance analysis were statistically significant, Tukey honestly significant differ-ence (HSD) or Kruskal-Wallis multiple comparison test was used to determine which group was different from others.
P values less than .05 were considered to be significant.
Results
Oxidative stress
The levels of MDA and GSH are summarized inTable 1. Both the liver levels of MDA and GSH were significantly different between the honey and the BDL groups (P⫽ .013 and .003, respectively). However, there was no significant difference between the plasma MDA and GSH levels of these groups (P⬎ .05).
Histopathological results
Tissues from group I presented no morphologic alter-ations in the portal tract.Fig. 1A shows a typical portal tract containing the terminal branch of the hepatic portal vein, terminal branches of the hepatic artery with the structure of arterioles, and bile ductules. The bile ducts are usually located at the periphery of the tract. The tissues of group II displayed some histopathologic changes in the portal tract such as proliferation of the duct epithelial cells and looping and reduplication of the ducts and ductules. The surround-ing hepatocytes were swollen and undergosurround-ing degeneration (Fig. 1B and D). In group III, the histopathologic evidence of the bile ductules’ proliferation was markedly reduced (Fig. 1C). The removal of the duct epithelial cells via apoptosis also supported this finding. The characteristic morphologic features of apoptosis, including nuclear frag-mentation and cellular shrinkage, were observed in electron microscopic sections. The apoptosis and degeneration of these epithelial cells would cause a reduction in the number of ducts in the portal tract (Fig. 1E).
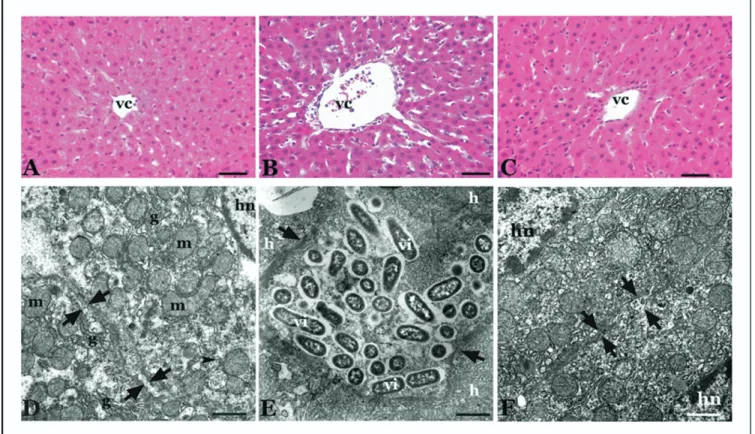
Group I did not show any difference from the normal lobul structure of liver tissue such as hepatocytes surround-ing the terminal hepatic venule (Fig. 2A). The hepatocyte cytoplasm is crowded with organels like rough endoplasmic reticulum and smooth endoplasmic reticulum, and rich with mitochondria. The plasma membranes of adjacent hepato-cytes especially close to canalicules are tightly bound and
Table 1
MDA and GSH levels of liver and plasma
Variable Sham BDL BDL⫹ honey P
MDA, liver (nmol/mg tissue) .4886⫾ .0613 .6335⫾ .1622† .4786⫾ .0757 .013* GSH, liver (nmol/mg tis.) .0612⫾ .0048 .0451⫾ .0055¶ .0629⫾ .0058 .003* MDA, plasma IU/mL 5.6993⫾ 1.5502‡ 9.6434⫾ 2.8760 8.5007⫾ 2.3820 .010* GSH, plasma IU/mL .5143⫾ .1263 .4358⫾ .1023 .5880⫾ .3224 .201*
Note: The liver levels of MDA and GSH were significantly different between group II and III while there was no difference in plasma levels. * One-Way ANOVA.
† BDL vs BDL⫹ honey was statistically significant (P ⫽ .030). ‡ Sham vs BDL was statistically significant (P⫽ .013). ¶ BDL vs BDL⫹ honey was statistically significant (P ⫽ .029).
the intercellular space width was normal (Fig. 2D). When we investigated the micropgraphs of group II, enlargement of hepatocytes with dilated canalicular spaces was seen. The enlarged hepatocytes seemed to be separated from each other because of the edema. Collagen deposition and edema around vena centralis were also observed (Fig. 2B). Numer-ous irregular mikrovilli extending from the hepatocyte sur-face were seen in the remarkable dilated bile canaliculi (Fig. 2E). In the sections of group III, we found conspicuous reduction in the size of enlarged hepatocytes and edema (Fig. 2C). The dilatation of the bile canaliculi between the 2 adjacent hepatocytes dramatically turned to original dimen-tion (Fig. 2F).
TUNEL assay
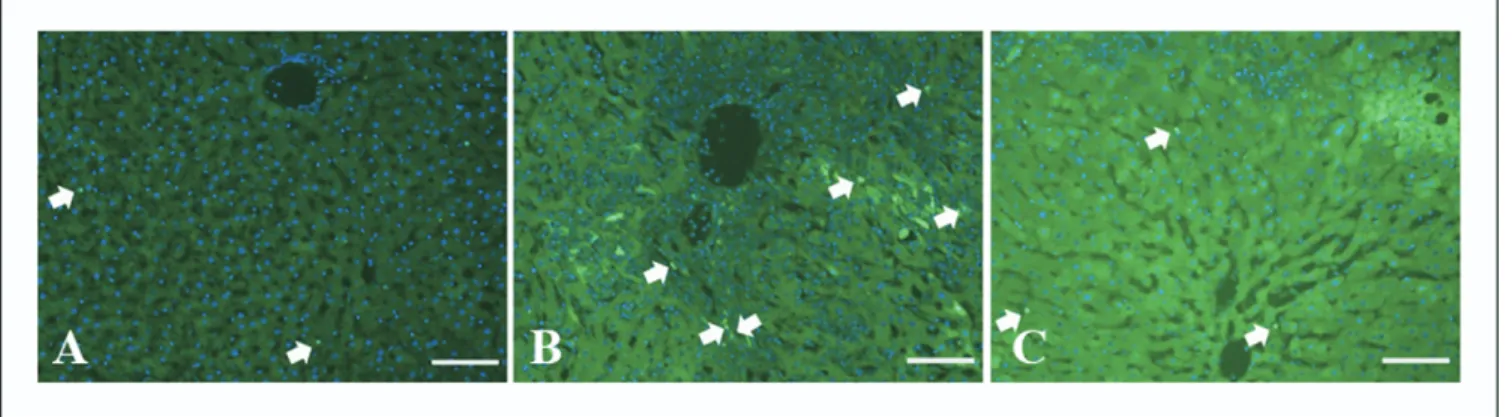
TUNEL assay results are listed inTable 2. There was a significant difference between the sham-operated and BDL groups (P⬍ .001). Administration of honey decreased the number of apoptotic cells compared with the BDL group (P⬍ .001) (Fig. 3).
Comments
The liver is a multifunctional organ that plays essential roles in metabolism, biosynthesis, excretion, secretion, and detoxification. Enhanced liver cell death and impaired re-generation are indeed features of most liver disorders. How-ever, unlike almost all other organs, the liver can regenerate from even massive cellular loss by cell proliferation. When cellular loss exceeds a certain proportion, regeneration fails, and hepatic failure and death occur[27]. Biliary obstruction causes 2 mechanical problems. First, substances excreted into the bile cannot enter the gastrointestinal tract, interrupt-ing the enterohepatic circulation. Second, obstruction of the extrahepatic biliary tree causes elevated pressure in the major ducts, which impairs the excretion of bile and may cause bile reflux. In addition to the changes in bilirubin and bile salt metabolism and the enterohepatic circulation, there are several alterations in other critical functions of the liver after biliary obstruction. Synthesis of proteins, including albumin, fibrinogen, prothrombin, haptoglobin, transferrin, ceruloplasmin, and clotting factors V, VII, IX, and X, is Fig. 1. Terminal branch of the hepatic portal vein (pv), terminal branch of the hepatic artery (a), bile ductules (*) are shown in the portal area of the liver biopsies in (A) group I, (B) group II (B), and (C) group III. Scale bar⫽ 20m. (D) The lumen of the bile ducts (L) and the proliferation of the bile duct epithelial cells (arrowheads) are seen in group II. Scale bar⫽ 9.3m. (E) Common morphology of hepatocyte nucleus (hn), still present bile pigment (bp) and the apoptotic bile duct epithelial cell (arrow) in group III. Scale bar⫽ 2.6m. (A, B, C) Mallory-Azan staining by light microscope; (D, E) TEM images.
impaired after BDL. The major complications of untreated obstructive jaundice include cholangitis, coagulation de-fects, and liver damage progressing to biliary fibrosis and cirrhosis[28].
In cholestatic liver injury, elevated concentrations of bile acids accumulate in the tissue and within the hepatocytes as a consequence of reduced bile flow, and trigger liver injury. Although the mechanisms of liver damage associated with cholestasis are complex and multifactorial, bile acid–medi-ated hepatotoxicity certainly plays a pivotal role in the pathogenesis of the disease. Hydrophobic bile acids have been shown to induce hepatic apoptosis in vitro and in vivo. Although apoptosis can be triggered by several different
stimuli, apoptotic signaling within the cell is transduced mainly via 2 molecular pathways: the death receptor path-way (extrinsic pathpath-way) and the mitochondrial pathpath-way (intrinsic pathway). The end point of both pathways is activation of a wide variety of intracellular proteases, espe-cially a group of proteolytic enzymes called caspases, and endonucleases that ultimately degrade the cellular constitu-ents. The intrinsic pathway is triggered by different extra- or intracellular signals such as␥-irradiation, oxidative stress, toxins, reactive intermediates of xenobiotic metabolism, growth factor deprivation, or some chemotherapeutic drugs that induce mitochondrial dysfunction [29]. Faubion et al [30]reported that glycochenodeoxycholate-induced hepato-cyte apoptosis involves ligand-independent oligomeriza-tion of Fas, recruitment of Fas-associated death domain (FADD), activation of caspase-8, and subsequent activation of effector proteases, including downstream caspases and cathepsin B.
Oxidative stress and lipid peroxidation are also involved in the pathogenesis of the cholestatic liver injury. Although Baron et al[31]concluded that the detergent action of bile salts was responsible for solubilization of plasma mem-branes and cell death, which in turn might lead to oxidative stress, glutathione oxidation, and lipid peroxidation, several studies have shown the role of oxygen free radicals and the protective effect of antioxidants in the cholestasis syndrome Fig. 2. The hepatocytes surrounding vena centralis (vc) in (A) group I, (B) group II, and (C) group III. Scale bar⫽ 10m. (D) The regular intercellular space between 2 hepatocytes (between the arrows), mitochondrion (m), hepatocyte nucleus (hn), and the smooth endoplasmic reticulum (g) are shown in group I. Scale bar⫽ 1.21m. (E) Hepatocytes (h), hepatocellular space (between the arrows), and numerous microvilli (vi) are shown in group II. Scale bar ⫽ 1.56m. (F) Hepatocyte nucleus (hn) and the regular intercellular space between the adjacent hepatocytes (between the arrows) are seen in group III. Scale bar⫽ 1.21m. (A, B, C) Hematoxylin and eosin staining by light microscope; (D, E, F) TEM images.
Table 2
The mean number of TUNEL-positive cells per field
Sham BDL BDL⫹ honey P
Mean number of TUNEL-positive
cells 1.10⫾ .73 5.50 ⫾ 1.07† 2.22 ⫾ .97†,‡ ⬍.001* Note: There was a significant difference between group I and II and/or group II and III.
* Kruskal-Wallis test.
† Difference between sham was statistically significant (P⬍ .001). ‡ Difference between BDL was statistically significant (P⬍ .001).
[32–39]. In these studies, antioxidant agents such as allo-purinol, melatonin, alpha tocopherol, ebselen, and vitamin E were shown to prevent cholestatic liver injury. Vendemiale et al [35] found that cholestasis was associated with a decreased protein and non-protein sulfhydryl content in the liver with an increased lipid peroxidation. They also showed that these alterations reversed almost completely after bili-ary drainage, indicating the cholestasis itself played a key role in the oxidative alterations found in the liver of patients affected by extrahepatic biliary obstruction. In another study, Ljubuncic et al[36]evaluated the extrahepatic tissue damage in cholestatic liver disease and found that experi-mental cholestasis was associated with increased lipid per-oxidation in the kidney, brain, and heart. According to these data, they concluded that oxidative stress in cholestatic liver disease was a systemic phenomenon probably influencing all tissues and organs, even those separated by the blood-brain barrier.
As we mentioned earlier, hydrophobic bile acids may cause hepatocellular apoptosis during cholestatic liver dis-ease. Sokol et al [39] investigated the mechanism of this injury, and determined the relationship of oxidant stress and the mitochondrial membrane permeability transition in he-patocyte necrosis induced by bile acids. They found that oxidant stress, not detergent effects or stimulation of cal-pain-like proteases, mediated the glycochenodeoxycholic acid–induced mitochondrial membrane permeability transi-tion in hepatocytes. They proposed that reducing mitochon-drial generation of reactive oxygen species or preventing increases in mitochondrial calcium might protect the hepa-tocyte against bile acid–induced necrosis. In another study, Yerushalmi et al[37]determined the roles of oxidant stress and the mitochondrial permeability transition in bile acid-induced apoptosis and concluded that reactive oxygen spe-cies generation, mitochondrial permeability transition, and cytochrome c release were critical steps in the induction of apoptosis by bile acids. They also found that antioxidants might reduce liver injury caused by low levels of bile acids by preventing the generation of oxidant stress.
Although these mechanisms seem to be responsible for liver injury, there may be other factors accompanying cho-lestasis in addition to bile acid cytotoxicity. Kupffer cell activation and increased neutrophil chemotaxis have been shown in obstructive jaundice models. However, it is not
clear whether the inflammatory cascades are primary or secondary[40].
Honey has bactericidal, bacteriostatic, antifungal, antivi-ral, scolicidal, antioxidant, antitumoantivi-ral, hepatoprotective, and anti-inflammatory effects [8 –22]. Al-Waili et al [20] assessed the hepatoprotective effects of commercial regular diet as control, total food restriction with honey, commer-cial regular diet with dextrose, or total food restriction with dextrose after carbon tetrachloride administration. During absolute honey feeding, elevations in alanine aminotrans-ferase (ALT) and aspartate aminotransaminotrans-ferase (AST) follow-ing carbon tetrachloride administration were significantly less than the values obtained in all other groups, with lower elevation in AST and ALT as compared to baseline values. Honey increased serum albumin, serum protein, and blood glucose, and caused a lower reduction in hemoglobin. The authors explained the influence of honey in carbon tetra-chloride–induced liver injury by its content of monosaccha-rides, its ability to inhibit prostaglandins, its ability to in-crease nitric oxide production, and its contents of antioxidants and trace elements. In another study, these authors also found that honey feeding during total food restriction significantly modified and ameliorated biochem-ical and hematologic changes observed after acute blood loss. They proposed to use honey as a part of bleeding management and during a food restriction regimen[21].
In our study, honey demonstrated a significant hepato-protective effect in an obstructive jaundice model. There was an evident reduction in the size of enlarged hepatocytes and the edema in the sections of group III compared with the BDL group (Fig. 2C). The dilatation of the bile canal-iculi between the 2 adjacent hepatocytes dramatically turned to original dimention (Fig. 2F). The antioxidant activity of honey has been evaluated in many studies[18,19,41– 45]. The composition of honey is rather variable and primarily depends on the floral source. Certain external factors such as seasonal and environmental factors also play a role. A wide range of minor constituents is also present in honey, many of which are known to have antioxidant properties. These include phenolic acids, flavonoids, ascorbic acid, carote-noid-like substances, organic acids, Maillard reaction prod-ucts, amino acids, proteins, and enzymes. The total antiox-idant capacity of honey is likely the result of the combined activity and interactions of these compounds[41]. Although Fig. 3. TUNEL assay of liver tissue. (A) Group 1, scale bar⫽ 40m; (B) group II, scale bar ⫽ 20 m; (C) group III, scale bar ⫽ 40 m. The blue staining is nucleus with Hoecht and the green reflection is the apoptotic nucleus (arrow) with TUNEL.
antimicrobial activity of honey is mainly attributed to hy-drogen peroxide, non-peroxide components (eg, antioxi-dants) may inhibit the growth of some food-borne patho-gens [45]. This antioxidant effect also plays a role in the wound-healing potential of honey[42].
The antioxidant effect of honey is not only local but also systemic. Little is known about the absorption of the phe-nolic acids and flavonoids that play a significant role in the antioxidant capacity of honey. In contrast to many other phenolics in foods or beverages such as tea, honey phenolics are present in the aglyconic rather than glycosidic form due to hydrolysis by the glycosidases present in the bee salivary glands. Phenolic aglycons are more readily absorbed through the gut barrier than their corresponding glycosides by passive diffusion and, therefore, the phenolics present in honey may be more readily bioavailable than the tea phe-nolics. In humans, serum antioxidant capacity increases 90 minutes after consumption of the buckwheat honey in water [18]. There is a linear correlation between phenolic content, oxygen radical absorbance capacity, and inhibition of li-poprotein oxidation of different honeys. Due to these prop-erties, honey may be used as a healthy alternative to sugar in many products and thereby serve as a source of dietary antioxidants[43]. Schramm et al[19]determined the effects of corn syrup and buckwheat honey on the plasma antiox-idant capacity in healthy human adults. They found that following consumption of honey treatment, plasma total phenolic content increased as did plasma antioxidant capac-ity. These data supported the concept that phenolic antioxi-dants from processed honey were bioavailable, and that they increased antioxidant activity of plasma. Authors speculated that these compounds might augment defenses against ox-idative stress and that these antioxidants might be able to protect humans from oxidative stress. Since the liver levels of MDA and GSH were significantly different in the honey group compared with the BDL group in our study, we concluded that the hepatoprotective effect of honey, at least in part, might be due to its antioxidant activity.
Honey may also protect liver damage in obstructive jaundice by its anti-inflammatory effect and by enhancing host immune defenses. It provides glucose supply for leu-cocytes, modulates the activation state of immunocompe-tent cells, affects the proliferative activity of human B and T lymphocytes and the activity of phagocytes, and stimu-lates human myeloid cell lines[10,17,45].
In the present study, we investigated the hepatoprotective effect of honey in obstructive jaundice model and found that honey diminished the negative effects of BDL on hepatic ultrastructure. We conclude that this effect of honey might be due to its antioxidant and anti-inflammatory activities, but further studies are needed to evaluate the exact mecha-nism of the hepatoprotective effect of honey.
References
[1] Zimmermann KC, Bonzon C, Green DR. The machinery of pro-grammed cell death. Pharm Ther 2001;92:57–70.
[2] Black DM, Behrns KE. Hepatic apoptosis and regeneration. Surg Oncol Clin North Am 2002;11:849 – 64.
[3] Bomzon A, Holt S, Moore K. Bile acids, oxidative stress, and renal function in biliary obstruction. Semin Nephrol 1997;17:549 – 62.
[4] Colette CP, Prosser CC, Yen RD, et al. Molecular therapy for hepatic injury and fibrosis: where are we? World J Gastroenterol 2006;12: 509 –15.
[5] Chuang JH, Chang NK, Huang CC, et al. Biliary intervention aug-ments chemotactic reaction and aggravates cholestatic liver injury in rats. J Surg Res 2004;120:210 – 8.
[6] Lechner AJ, Velasquez A, Knudsen KR, et al. Choleastatic liver injury increases circulating TNF-␣ and IL-6 and mortality after
Escherichia coli endotoxemia. Am J Respir Crit Care Med 1998;157:
1550 – 8.
[7] Ding JW, Andersson R, Soltesz V, et al. Obstructive jaundice impairs reticuloendothelial function and promotes bacterial translocation in the rat. J Surg Res 1994;57:238 – 45.
[8] Crane E. The past and present importance of bee products to man. In: Mizrahi A, Lensky Y, editors. Bee Products. New York: Plenum; 1997:1–13.
[9] Schmidt JO. Bee products: Chemical composition and application. In: Mizrahi A, Lensky Y, editors. Bee Products. New York: Plenum; 1997:15–26.
[10] Lusby PE, Coombes A, Wilkinson JM. Honey: a potent agent for wound healing? J Wound Ostomy and Continence Nursing 2002;29: 295–300.
[11] Lusby PE, Coombes AL, Wilkinson JM. Bactericidal activity of different honeys against pathogenic bacteria. Arch Med Res 2005;36: 464 –7.
[12] Cooper RA, Molan PC, Harding KG. The sensitivity to honey of gram-positive cocci of clinical significance isolated from wounds. J Appl Microbiol 2002;93:857– 63.
[13] Cooper RA, Halas E, Molan PC. The efficacy of honey in inhibiting strains of Pseudomonas aeruginosa from infected burns. J Burn Care Rehabil 2002;23:366 –70.
[14] Irish J, Carter DA, Shokohi T, et al. Honey has an antifungal effect against Candida species. Med Mycol 2006;44:289 –91.
[15] Al-Waili NS. Topical honey application vs. acyclovir for the treat-ment of recurrent herpes simplex lesions. Med Sci Monit 2004;10: MT94 – 8.
[16] Kilicoglu B, Kismet K, Koru O, et al. The scolicidal effects of honey. Adv Ther 2006;23:1085–91.
[16] Abuharfeil N, AlOran R, Aboshehada M. The effect of bee honey on the proliferative activity of human B and T-lymphocytes and the activity of phagocytes. Food Agricult Immunol 1999;11:169 –77. [17] Gheldof N, Wang XH, Engeseth NJ. Buckwheat honey increases
serum antioxidant capacity in humans. J Agric Food Chem 2003;51: 1500 –5.
[18] Schramm DD, Karım M, Schrader HR, et al. Honey with high levels of antioxidants can provide protection to healthy human subjects. J Agric Food Chem 2003;51:1732–5.
[19] Al-Waili NS, Saloom KY, Al-Waili TN, et al. Influence of various diet regimens on deterioration of hepatic function and hematological parameters following carbon tetrachloride: a potential protective role of natural honey. Nat Prod Res 2006;20:1258 – 64.
[20] Al-Waili N, Saloom KY, Akmal M, et al. Honey ameliorates influ-ence of hemorrhage and food restriction on renal and hepatic func-tions, and hematological and biochemical variables. Int J Food Sci Nutr 2006;57:353– 62.
[21] Mabrouk GM, Moselhy SS, Zohny SF, et al. Inhibition of methylni-trosourea (MNU) induced oxidative stress and carcinogenesis by orally administered bee honey and Nigella grains in Sprague Dawely rats. J Exp Clin Cancer Res 2002;21:341– 6.
[22] Kavutcu M, Canbolat O, Ozturk S, et al. Reduced enzymatic antiox-idant defense mechanism in kidney tissues from gentamicin-treated guinea pigs: effects of vitamins E and C. Nephron 1996;72:269 –74. [23] Lowry OH, Rosebrough NJ, Farr AL, et al. Protein measurement with
the Folin phenol reagent. J Biol Chem 1951;193:265–75.
[24] Dahle LK, Hill EG, Holman RT. The thiobarbituric acid reaction and the autoxidations of polyunsaturated fatty acid methyl esters. Arch Biochem Biophys 1962;98:253– 61.
[25] Paglia DE, Valentine WN. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med 1967;70:158 – 69.
[26] Malhi H, Gores GJ, Lemasters JJ. Apoptosis and necrosis in the liver: a tale of two deaths? Hepatology 2006;43:S31– 44.
[27] Scott-Conner CEH, Grogan JB. The pathophysiology of biliary ob-struction and its effect on phagocytic and immune function. J Surg Res 1994;57:316 –36.
[28] Guicciardi ME, Gores GJ. Apoptosis: a mechanism of acute and chronic liver injury. Gut 2005;54:1024 –33.
[29] Faubion WA, Guicciardi ME, Miyoshi H, et al. Toxic bile salts induce rodent hepatocyte apoptosis via direct activation of Fas. J Clin Invest 1999;103:137– 45.
[30] Baron V, Muriel P. Role of glutathione, lipid peroxidation and anti-oxidants on acute bile duct obstruction in the rat. Biochem Biophy Acta 1999;1472:173– 80.
[31] Mun KC, Kwak CS, Kwon KY. The protective effect of allopurinol on cholestatic liver injury induced by bile duct ligation. J Korean Med Sci 1996;11:239 – 43.
[32] Ohta Y, Imai Y, Matsura T, et al. Successively postadministered melatonin prevents disruption of hepatic antioxidant status in rats with bile duct ligation. J Pineal Res 2005;39:367–74.
[33] Ara C, Kirimlioglu H, Karabulut AB, et al. Protective effect of resveratrol against oxidative stress in cholestasis. J Surg Res 2005; 127:112–7.
[34] Vendemiale G, Grattagliano I, Lupo L, et al. Hepatic oxidative alterations in patients with extra-hepatic cholestasis. Effect of surgical drainage. J Hepatol 2002;37:601–5.
[35] Ljubuncic P, Tanne Z, Bomzon A. Evidence of a systemic phenomenon for oxidative stress in cholestatic liver disease. Gut 2000;47:710 – 6. [36] Yerushalmi B, Dahl R, Deverequx MW, et al. Bile acid-induced rat
hepatocyte apoptosis is inhibited by antioxidants and blockers of the mitochondrial permeability transition. Hepatology 2001;33:616 –26.
[37] Montilla P, Cruz A, Padillo FJ, et al. Melatonin versus vitamin E as protective treatment against oxidative stress after extra-hepatic bile duct ligation in rats. J Pineal Res 2001;31:138 – 44.
[38] Sokol RJ, Straka MS, Dahl R, et al. Role of oxidant stress in the permeability transition induced in rat hepatic mitochondria by hydro-phobic bile acids. Pediatr Res 2001;49:519 –31.
[39] Guicciardi ME, Gores GJ. Cholestatic hepatocellular injury: what do we know and how should we proceed? J Hepatol 2005;42:297–300. [40] Gheldof N, Wang XH, Engeseth NJ. Identification and quantification of antioxidant components of honeys from various floral sources. J Agric Food Chem 2002;50:5870 –7.
[41] Henriques A, Jackson S, Cooper R, et al. Free radical production and quenching in honeys with wound healing potential. J Antimicr Che-mother 2006;58:773–7.
[42] Gheldof N, Engeseth NJ. Antioxidant capacity of honeys from vari-ous floral sources based on the determination of oxygen radical absorbance capacity and inhibition of in vitro lipoprotein oxidation in human serum samples. J Agric Food Chem 2002;50:3050 –5. [43] Bilsel Y, Bugra D, Yamaner S, et al. Could honey have a place in
colitis therapy? Effects of honey, prednisolone, and disulfiram on inflammation, nitric oxide, and free radical formation. Dig Surg 2002;19:306 –12.
[44] Taormina PJ, Niemira BA, Beuchat LR. Inhibitory activity of honey against foodborne pathogens as influenced by the presence of hydro-gen peroxide and level of antioxidant power. Int J Food Microbiol 2001;69:217–25.
[45] Watanabe K, Shinmoto H, Kobori M, et al. Stimulation of cell growth in the U-937 human myeloid cell line by honey royal jelly protein. Cytotechnology 1998;26:23–7.