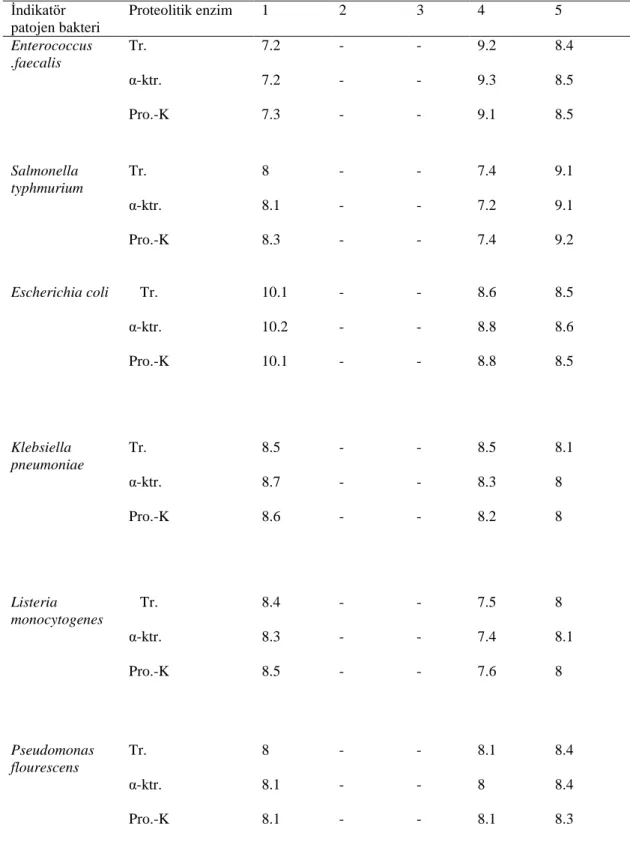
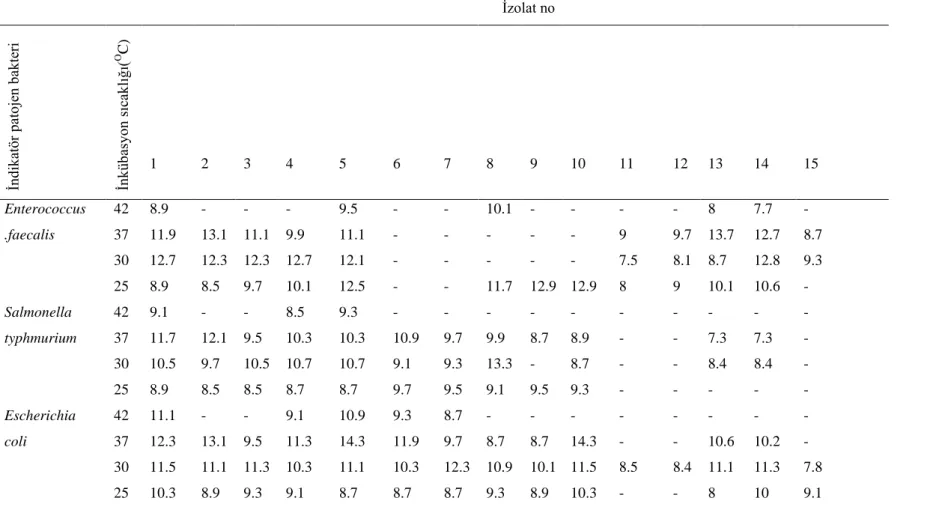
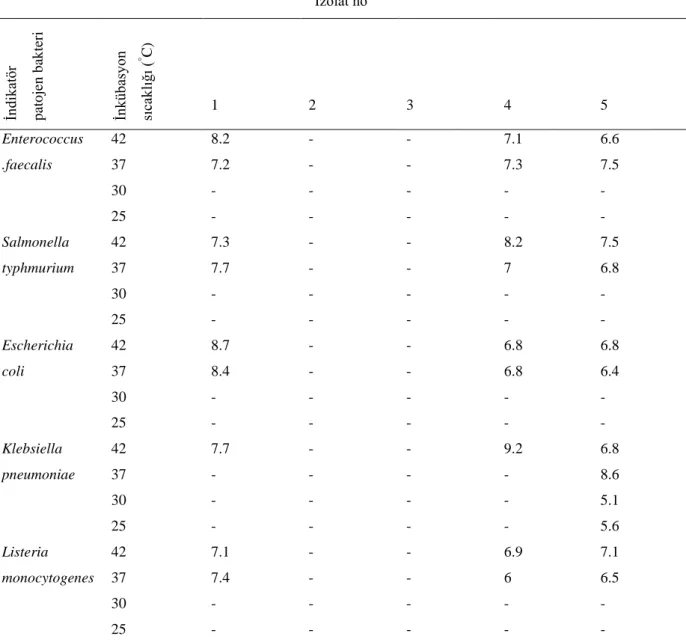
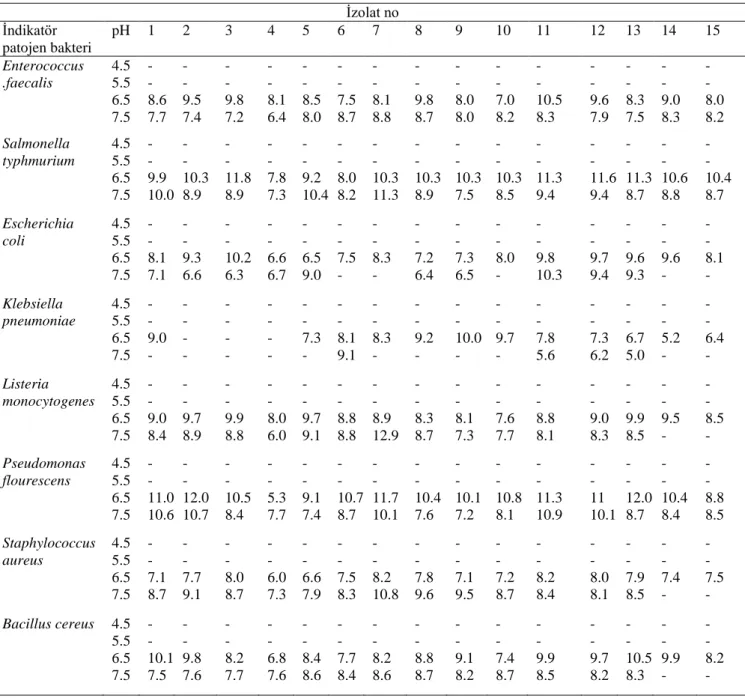
Doğal olarak üretilen yoğurtlardan izole edilen Streptococcus thermophilus ve Lactobacillus delburieckii subsp bulgaricus'un bakteriyosin üretme yeteneklerinin belirlenmesi ve ürettikleri bakteriyosinlerin karakterizasyonu
87
0
0
Tam metin
Şekil
+6

Benzer Belgeler
S.No Ders Dersin Adı Hs Yer Dersin Öğretmenleri..
Bir üniteyi kapsayan, orta ve ileri düzey sorulardan oluşan..
2 SBYLJ SEÇMELİ BİYOLOJİ 4 MESUT DEMİR. 3 SFZK SEÇMELİ FİZİK 4
ÇANKAYA BAHÇELİEVLER 100YIL MESLEKİ VE TEKNİK ANADOLU LİSESİ..
15 TDED TÜRK DİLİ VE EDEBİYATI 5 ARZU TOPRAK. 16 YDL2 YABANCI DİL 2
SİNCAN YUNUS EMRE ANADOLU İMAM HATİP LİSESİ.. S.No Ders Dersin Adı Hs Yer
8 SMEK SEÇMELİ MEKANİZMALAR 2 MEHMET BODUR ERCAN ERTEN. 205 9 SİŞETD SEÇMELİ İŞ ETÜDÜ
2 BYD4 BİRİNCİ YABANCI DİL 4 EMİNE ÖZBEN ULUTÜRK. 3 BYS27 BİYOLOJİ (SEÇMELİ-4) 4
