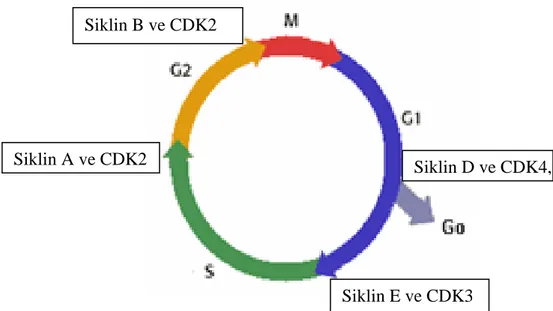
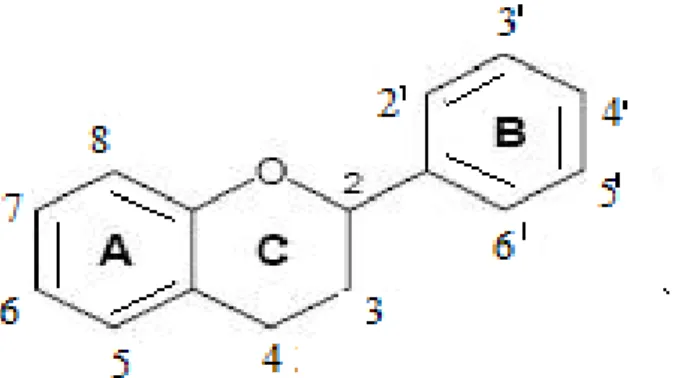
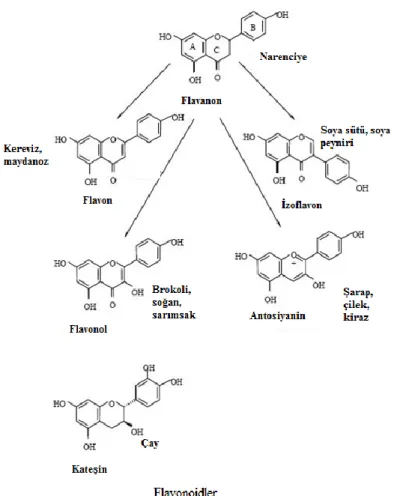
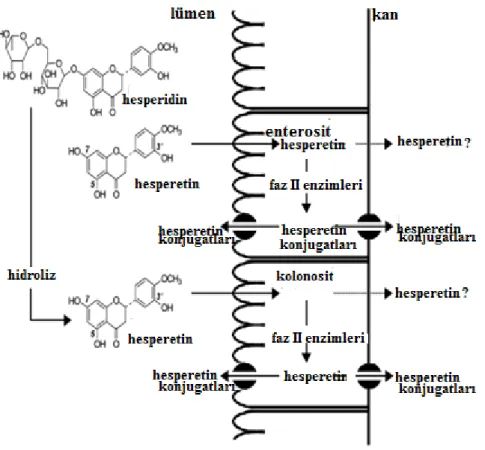
Flavonoidlerin kanser hücrelerine etkisi
Tam metin
Şekil

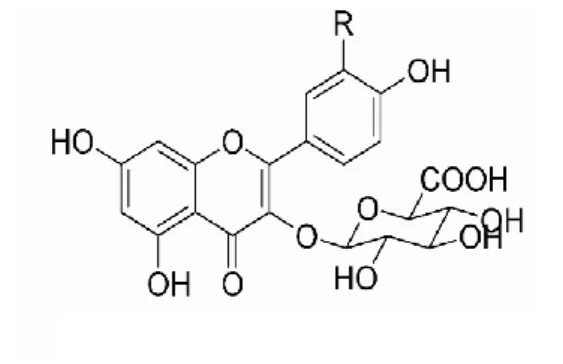
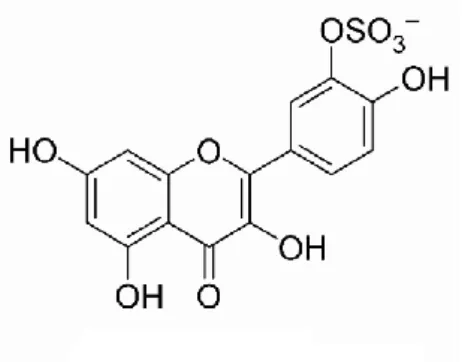
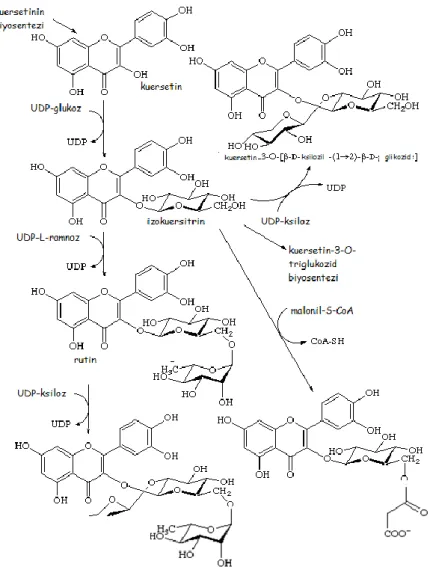

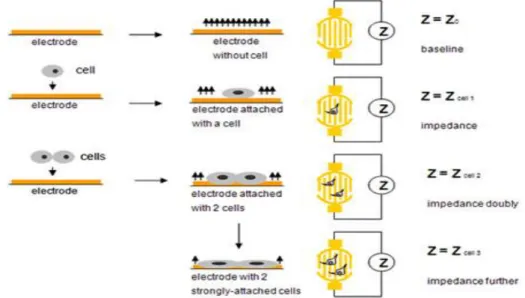
Benzer Belgeler
Duygusal zekânın alt boyutu olan kendi duygularını değerlendirme boyutu yine diğer alt boyutlar olan başkalarının duygularını değerlendirme (r=0,404) duyguların
Bu değerlendirmeler üçgen bulanık sayılara dönüştürülerek, bulanık TOPSIS yöntemi çalıştırılmış ve sonuca göre ilk sıradaki tedarikçi, işletme için en
Türkiye’nin çok geniş bir çerçevede değişen büyüme oranları bu çalışmada etki edilen sistem değişkeni olarak ele alınmıştır. Büyüme oranına etki eden en
Çevresel Psikoloji'de "çevre", birey dışındaki herşey olarak tanımlanıyor. Bu tanım, hem doğal ve yapılı çevreyi, hem de toplumsal çevreyi
Sonuç olarak taşra siyaseti; merkezle olan ilişkileri, giderek demokratikleşen yapısı, siyasal aktörleri, birincil ve cemaatsel ilişkileri, köklü aileleri,
Türkiye’de ve dünyada faaliyet gösteren, beyaz eşya sektörünün önde gelen işletmelerin tedarik zincirinde kurumsal sosyal sorumluluk faaliyetleri ve bu faaliyetler
sınıf Sosyal Bilgiler dersi Türk Tarihinde Yolculuk ünitesi öğretiminde internet tabanlı öğretim yönteminin öğrencilerin akademik baĢarılarına etkisi ve hatırda
Ayrıca n kodun uzunluğu, m otomorfizmanın mertebesi olmak üzere (n,m)=1 olması durumunda skew cyclic kodun cyclic koda eşit, (n,m)=d olması durumunda quasi-
