ANTİOKSİDAN ENZİMLERİN VE GLOBAL PROTEİN FOSFORİLASYONUNUN DİYABET VE ANTİOKSİDAN UYGULAMALARI İLE DEĞİŞİMLERİNİN ARAŞTIRILMASI
Davut BOZAN Yüksek Lisans Tezi Biyoloji Anabilim Dalı
Moleküler Biyoloji ve Genetik Programı Yrd. Doç. Dr. Gökhan SADİ
T.C.
KARAMANOĞLU MEHMETBEY ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
ANTİOKSİDAN ENZİMLERİN VE GLOBAL PROTEİN
FOSFORİLASYONUNUN DİYABET VE ANTİOKSİDAN UYGULAMALARI İLE DEĞİŞİMLERİNİN ARAŞTIRILMASI
YÜKSEK LİSANS TEZİ Davut BOZAN
Anabilim Dalı: Biyoloji
Programı : Moleküler Biyoloji ve Genetik
Tez Danışmanı: Yrd. Doç. Dr. Gökhan SADİ
TEZ BİLDİRİMİ
Tez yazım kurallarına uygun olarak hazırlanan bu tezin yazılmasında bilimsel ahlak kurallarına uyulduğunu, başkalarının eserlerinden yararlanılması durumunda bilimsel normlara uygun olarak atıfta bulunulduğunu, tezin içerdiği yenilik ve sonuçların başka bir yerden alınmadığını, kullanılan verilerde herhangi bir tahrifat yapılmadığını, tezin herhangi bir kısmının bu üniversite veya başka bir üniversitedeki başka bir tez çalışması olarak sunulmadığını beyan ederim.
ÖZET Yüksek Lisans Tezi
ANTİOKSİDAN ENZİMLERİN VE GLOBAL PROTEİN
FOSFORİLASYONUNUN DİYABET VE ANTİOKSİDAN UYGULAMALARI İLE DEĞİŞİMLERİNİN ARAŞTIRILMASI
Davut BOZAN
Karamanoğlu Mehmetbey Üniversitesi Fen Bilimleri Enstitüsü
Biyoloji Anabilim Dalı
Danışman: Yrd. Doç. Dr. Gökhan SADİ Haziran, 2013, 73 sayfa
Diyabet vücutta birçok organı etkileyen ve bir dizi komplikasyon serisi olarak nitelendirilen bir hastalıktır. Diyabet sıçanlarda streptozotosinin tek doz uygulaması ile oluşturulmuş, resveratrol (RSV) ve C vitamini (VC) 50 mg/kg olacak şekilde sıçanlara intraperitonal yoldan günlük olarak üç hafta süre ile verilmiştir. Sıçanların karaciğer dokularında toplam fosfoprotein saflaştırması gerçekleştirilmiş ve fosfoprotein profilinin diyabet ve antioksidan uygulamaları ile değiştiği SDS-PAGE ile ortaya çıkarılmış, diyabet ve antioksidan uygulamalarının, dokulardaki toplam protein fosforlanma düzeylerini arttırıcı etkileri belirlenmiştir. Katalaz protein ifadelenmesinin diyabet uygulaması ile (p<0,05) düştüğü, RSV ve VC gibi iki güçlü antioksidanın bu düşüklüğü kontrol düzeyine yönlendirdiği belirlenmiştir. Diyabette fosfo KAT düzeyinin anlamlı derecede azaldığı, RSV ve VC’nin KAT fosforilasyon düzeyini daha da düşürdüğü ortaya çıkarılmıştır. Diyabet, karaciğer dokularındaki (unfosfo ve fosfo) glutatyon peroksidaz düzeylerinde anlamlı bir değişiklik oluşturmamış aynı şekilde antioksidanların doku fosforile ve forforile olmayan GPx düzeyini etkilemediği ifade edilmiştir. Real time PCR çalışmaları, KAT ve GPx enzimlerinin mRNA ekspresyonlarının diyabetik grupta kontrol grubuna göre anlamlı bir şekilde azaldığını (p<0,05) göstermiştir. RSV diyabetik sıçanlara uygulandığında karaciğer KAT ve GPx mRNA ekspresyonunu kontrol grubuna doğru yönlendirirken, VC istatistiksel olarak anlamlı bir etki göstermemiştir. Bu sonuçlara göre RSV antioksidan enzimler üzerine
etkilerini transkripsiyon düzeyinde gösterirken VC etkilerini post-translasyonel olarak ortaya çıkarmıştır.
Sonuç olarak bu çalışma ile diyabetin neden olduğu protein fosforilasyonu düzeyindeki değişiklikler ve bu değişiklikler üzerine RSV ve VC’nin etkileri ortaya çıkarılmış, aktivitelerinin fosforilasyon ile kontrol edildiği daha önceden bilinen KAT ve GPx enzimlerinin diyabette transkripsiyonel ve post-translasyonel olarak nasıl kontrol edildiklerine dair bilimsel veriler elde edilmiştir.
Anahtar Kelimeler: Diyabet, Oksidatif Stres, Antioksidan Enzimler, Protein Fosforilasyonu
ABSTRACT MSc. Thesis
INVESTIGATION OF CHANGES IN GLOBAL AND ANTIOXIDANT ENZYMES’ PROTEIN PHOSPHORYLATION BY DIABETES AND
ANTIOXIDANT TREATMENTS Davut BOZAN
Karamanoğlu Mehmetbey University Graduate School of Natural and Applied Sciences
Department of Biology
Supervisor: Asst. Prof. Dr. Gökhan SADİ June, 2013, 73 pages
Diabetes is a disease characterized as a series of complications which affects many organs in the body. Diabetes was induced in rats by single dose of streptozotocin injection; resveratrol (RSV) and vitamin C (VC) were given as 50 mg/kg in each day for three weeks by intraperitonal injection. After antioxidant administration, total phosphoprotein purification was carried out by metal affinity chromatography from liver tissues and modulation of phosphoprotein profiles of liver tissues were demonstrated using SDS-PAGE. Furthermore, augmentation in the total phosphoprotein amounts with diabetes and antioxidants were revealed. Catalase protein expressions were decreased (p<0,05), RSV and VC elevated this reduction towards the control levels. Besides, phospho CAT levels were also decreased in diabetics and antioxidant treatments further reduced this decrement. Diabetes and antioxidants did not change the unphospho and phospho glutathione peroxidase protein levels. Real Time PCR studies demonstrated that mRNA expressions of CAT and GPx enzymes were statistically (p<0,05) downregulated with diabetes as compared to control groups. As administered to the diabetic animals, RSV increased the gene expression levels of both antioxidant enzymes toward the controls but VC did not. According to these results, RSV exerts its effects on antioxidant enzymes at transcriptional level but VC has post-translational effect mainly via protein phosphorylation.
As a conclusion, changes in protein phosphorylation levels caused by diabetes and effects of two strong antioxidants RSV and VC on protein phosphorylation status were determined. Furthermore, the enzymes; CAT and GPx whose activities are known to be
regulated by phosphorylation were studied and their post-regulation mechanism by diabetes and antioxidant treatment were revealed with this study.
ÖNSÖZ
Öncelikle bana bu önemli araştırma konusunu veren, laboratuar çalışmalarım ve tezimin yazımı sırasında her türlü desteğini ve yardımını esirgemeyen ve beni bilgisiyle donatan değerli hocam sayın Yrd. Doç. Dr. Gökhan SADİ’ye tüm içtenliğimle teşekkürlerimi sunarım.
Ayrıca benden yardımlarını esirgemeyen değerli hocalarım Doç. Dr. Muhammad AASIM’a, Yrd. Doç. Dr. Aytaç KOCABAŞ’a ve Yrd. Doç. Dr. Mehmet Fatih AYDIN’a sonsuz teşekkürlerimi sunarım.
Yüksek lisans eğitimim boyunca beraber çok güzel vakit geçirdiğim ve tez çalışmalarımda yardımlarını esirgemeyen değerli dostlarım Arş. Gör. Buğrahan EMSEN’e, Arş. Gör. Osman AĞAR’a, Arş. Gör. Muhammet DOĞAN’a ve Arş. Gör. Musa KAMACI’ya çok teşekkür ederim.
Tüm hayatım boyunca olduğu gibi yüksek lisans eğitimimde de maddi manevi her türlü yönden beni destekleyen, bana sabır gösterip sıkıntılarımda dualarıyla yanımda olan annem ve babam, Şemse-Mustafa BOZAN başta olmak üzere, çok değerli kardeşim Şahin BOZAN’a ve tüm aileme şükranlarımı sunarım.
Son olarak değerli dostum Halis EKMEKCİ’ye ve laboratuarda beraber çalıştığım bütün araştırmacı arkadaşlarıma çok teşekkür ederim.
Davut BOZAN Haziran, 2013
İÇİNDEKİLER
Sayfa
ÖZET ... i
ABSTRACT ... iii
ÇİZELGELER DİZİNİ ... viii
SİMGELER VE KISALTMALAR DİZİNİ ... xii
1. GİRİŞ ... 1
2. KURAMSAL TEMELLER ve KAYNAK ARAŞTIRMASI ... 3
2.1. Serbest Radikaller ... 3
2.1.1. Serbest Radikal Çeşitleri ... 6
2.1.2. Serbest Radikallerin Etkileri ... 10
2.2. Vücudun Antioksidan Savunma Mekanizmaları... 16
2.2.1. Enzimatik Antioksidanlar ... 19
2.2.2. Enzimatik Olmayan Antioksidanlar ... 23
2.3. Oksidatif Stres ... 27
2.4. Diyabet ... 28
2.4.1. Deneysel Diyabetik Hayvan Modelleri ... 29
2.4.2. Diyabet ve Oksidatif Stres ... 30
2.5. Protein Fosforilasyonu ... 32
3. MATERYAL VE YÖNTEM ... 35
3.1. Materyal ... 35
3.1.1. Kullanılan Kitler ve Kimyasal Maddeler ... 35
3.2. Deneysel Diyabetin Oluşturulması ve Antioksidan Uygulaması ... 35
3.3. Toplam Fosfoprotein Saflaştırılması ... 36
3.4. Toplam RNA İzolasyonu, cDNA Sentezi ve Real Time PCR ile KAT ve GPx Gen Ekspresyonlarının Ölçülmesi... 47
4. BULGULAR ... 50
4.1. Deney Hayvanlarının Ağırlık ve Açlık Kan Şekeri Düzeylerinin Değişimleri .. 50
4.2. Fosfoprotein Fraksiyonlarının Protein Konsantrasyonları ... 51
4.3. Unfosfo Protein (Flow out) Fraksiyonunun Protein Konsantrasyonları ... 53
4.5. Glutatyon Peroksidaz Enziminin Western Blot Analiz Sonuçları ... 56
4.6. KAT ve GPx Enzimlerinin Gen Ekspresyonları ... 58
5. TARTIŞMA VE SONUÇ ... 59
6. KAYNAKLAR ... 63
ÇİZELGELER DİZİNİ
Çizelge Sayfa
Çizelge 2.1. Serbest oksijen radikalleri ... 4
Çizelge 2.2. Hücredeki serbest oksijen radikali kaynakları ... 6
Çizelge 2.3. Antioksidan savunma sistemleri ... 17
Çizelge 3.1. KAT, GPx ve β-aktin genleri için primer dizilimleri ... 48
Çizelge 4.1: Konsantre edilmemiş fosfoprotein fraksiyonlarının protein konsantrasyonları ... 51
Çizelge 4.2: Ultrafiltration kolonları kullanılarak konsantre edilen fosfoprotein fraksiyonlarının protein konsantrasyonları ... 53
Çizelge 4.3: Fosforlanmamış protein fraksiyonlarının konsantrasyonları ... 53
Çizelge 4.4: KAT ve GPx enzimlerinin kontrol, diyabetik, RSV verilmiş diyabetik (D+RSV) ve VC verilmiş diyabetik (D+VC) dokularda ölçülen mRNA ekspresyon miktarları ... 58
ŞEKİLLER DİZİNİ
Şekil Sayfa
Şekil 2.1. Mitokondriyal elektron iletim zinciri tarafından süperoksitin üretimi ... 7
Şekil 2.2. Hidrojen peroksitin moleküler şekli ... 8
Şekil 2.3. Ksantin oksidaz (XO) ile H2O2 üretimi ... 9
Şekil 2.4. Hücrede serbest radikallerin çeşitli hedefleri ... 10
Şekil 2.5. Serbest radikaller tarafından oluşabilen proteinler üzerindeki karbonilasyon ürünleri ... 11
Şekil 2.6. Hidroksil radikalinin pürin ve pirimidin bazları üzerine etkisiyle oluşan ürünler ... 13
Şekil 2.7. Serbest oksijen radikallerin oluşumu ve lipitlere etkileri ... 14
Şekil 2.8. Lipit peroksidasyonunun kimyasal mekanizması ... 15
Şekil 2.9. Serbest radikallerin oluşumu ve antioksidan enzimlerin etkisi ... 18
Şekil 2.10. Antioksidan enzimlerin glutatyon redoks döngüsü ile beraber serbest radikal kaynaklarını ortadan kaldırma mekanizması ... 20
Şekil 2.11. Antioksidan enzimler ve glutatyonun indirgenme reaksiyonu... 22
Şekil 2.12. Glutatyonun molekül şekli ... 23
Şekil 2.13. E vitamininin ve değişik fromlarının molekül şekli ... 24
Şekil 2.14: C vitamininin molekül şekli ... 25
Şekil 2.15: A vitamininin molekül şekli ... 26
Şekil 2.16: Trans-Resveratrol’ün molekül şekli ... 26
Şekil 2.17. Reaktif oksijen türevleri ve antioksidan mekanizmaları ... 29
Şekil 2.18. Serbest radikal zararı sonucu ortaya çıkan ürünler... 32
Şekil 2.19. Oksidatif NF-κB aktivasyonu için oluşturulmuş model ... 33
Şekil 2.20. Reseptör tirozin kinaz (RTK) ve tirozin fosforilasyonun modülatörü olan reaktif oksijen türleri (H2O2). ... 34
Şekil 3.1. Diyabetin hayvansal modelinin oluşturulması ve antioksidanların uygulaması ... 36
Şekil 3.2. Doku fosfoproteinlerinin saflaştırılmasının basamakları... 37
Şekil 3.3. Fosfoprotein saflaştırma kiti ile doku fosfoproteinlerinin saflaştırılması işlemi ... 38
Şekil 3.4. Thermo ScanGO Mikroplaka okuyucu ... 39
Şekil 3.5. Sıkıştırıcı ve ayrıştırıcı jellerin hazırlanışı ... 40
Şekil 3.6. Örneklerin jel kuyucuklarına yükleme işlemi ... 40
Şekil 3.7. SDS-PAGE, örneklerin yürütülmesi ve proteinlerin büyüklüklerine göre ayrıştırılması işlemi ... 41
Şekil 3.8. BioRad ChemiDocMP görüntüleme sistemi... 42
Şekil 3.9. Fosfoprotein saflaştırma kolonlarından çıkan örneklerin SDS-PAGE ardından gümüş boyama sonrası görüntüsü………... 42
Şekil 3.10. Seyreltik proteinlerin mikrospin ultrafiltrasyon kolonları ile konsantre edilmeleri ... 43
Şekil 3.11. Western Blot yönteminde transfer kasetlerinin hazırlanışı ve transfer edilişi ... 44
Şekil 3.12. Western-blot ile protein ifadelenmelerinin ölçüm yöntemi ... 45
Şekil 3.13. PVDF membranların antikorlar ile çalkalayıcıda inkübasyonu ... 46
Şekil 3.14. Kontrol, Diyabetik, D+RSV ve D+VC gruplarının ham homejenat, fosforlanmamış protein fraksiyonu ve fosfoprotein fraksiyonu kullanılarak gerçekleştirilen KAT ve GPx proteininin Western Blot sonuçları ... 47
Şekil 3.15. İzole edilen toplam RNA’ların agaroz jel elektroforez görüntüsü ... 48
Şekil 3.16. Roche LightCycler 480 II Real Time PCR cihazı ... 49
Şekil 4.1. Sıçan ağırlıklarının (gr) haftalara göre değişimleri ... 50
Şekil 4.2. Deney sürecinde hayvanların açlık kan şekeri (mg/dL) değişimleri ... 51
Şekil 4.3. Kontrol, diyabetik, RSV verilmiş diyabetik (D+RSV) ve VC verilmiş diyabetik (D+VC) sıçan karaciğer dokularında konsantre edilmemiş fosfoprotein fraksiyonlarının protein konsantrasyonları ... 52
Şekil 4.4. Kontrol, diyabetik, RSV verilmiş diyabetik (D+RSV) ve VC verilmiş diyabetik (D+VC) sıçan karaciğerlerinde ölçülen KAT proteinin western blot analiz sonuçları ... 54 Şekil 4.5. Kontrol, diyabetik, RSV verilmiş diyabetik (D+RSV) ve VC verilmiş diyabetik (D+VC) sıçan karaciğer dokularında toplam KAT protein ekspresyonu
Şekil 4.6. Kontrol, diyabetik, RSV verilmiş diyabetik (D+RSV) ve VC verilmiş diyabetik (D+VC) sıçan karaciğer dokularında fosfo KAT düzeyleri ... 56 Şekil 4.7. Kontrol, Diyabetik, RSV verilmiş diyabetik (D+RSV) ve VC verilmiş diyabetik (D+VC) sıçan karaciğerlerinde ölçülen GPx proteinin western blot analiz sonuçları ... 56 Şekil 4.8. Kontrol, diyabetik, RSV verilmiş diyabetik (D+RSV) ve VC verilmiş diyabetik (D+VC) sıçan karaciğer dokularında GPx düzeyleri ... 57 Şekil 4.9. Kontrol, diyabetik, RSV verilmiş diyabetik (D+RSV) ve VC verilmiş diyabetik (D+VC) sıçan karaciğer dokularında fosfo GPx düzeyleri ... 57
SİMGELER VE KISALTMALAR DİZİNİ Simgeler Açıklaması dH2O Distile Su g Gram H2O2 Hidrojen peroksit L Litre M Molar mg Miligram mL Mililitre mM Milimolar mmol Milimol 1 O2 Singlet oksijen O2. - Süperoksit radikali OH . Hidroksil radikali
R . Karbon merkezli radikaller
Kısaltmalar Açıklaması
BSA Bovin serum albumin
KAT Katalaz
Cu/Zn SOD Bakır/çinko süperoksit dismutaz
DNA Deoksiribo nükleik asit
EDTA Etilendiamin tetraasetikasit
FAD Flavin adenin dinükleotit
FCR Folin-Ciocalteu reaktifi
Fe SOD Demir süperoksit dismutaz
GSH Glutatyon
GPx Glutatyon peroksidaz
GR Glutatyon redüktaz
GST Glutatyon-S-transferaz
LOOH Lipit hidroperoksit
LPO Lipit peroksidasyonu
L(R)OOH Hidroperoksit
MDA Malondialdehit
Mn SOD Mangan süperoksit dismutaz
NAD Nikotinamid adenin dinükleotid
NAD+ Yükseltgenmiş nikotinamid adenin
NADH İndirgenmiş nikotinamid adenin
NADPH Nikotinamid adenindinükleotid fosfat
NBT Nitroblue tetrazolyum
NO Nitrik oksit
RNA Ribonükleik asit
ROO Peroksil radikali
ROS (ROT) Reaktif oksijen türleri
RSV Resveratrol
SOD Süperoksit dismutaz
UV Ultraviyole
VC C vitamini
1. GİRİŞ
Serbest radikaller yapılarında eşlenmemiş elektron bulunduran ve çevresindeki her madde ile tepkime verme yatkınlığı olan, negatif yüklü elektron sayısının çekirdekteki pozitif yüklü proton sayısı ile eşit olmadığı moleküllerdir (Fang ve ark., 2002). Serbest radikaller; yapıları, fiziksel ve kimyasal özellikleri, hücresel kaynakları, rol oynadıkları tepkimeler ve etkileri ile çeşitli klinik durumların patojenezinde rol oynarlar (Wassmann ve ark., 2004).
Yaşamsal faaliyetlerini sürdürebilmeleri için aerobik organizmalar tarafından organik moleküllerden enerji elde edilmesinde oksijenin kullanılması, bu organizmalarda toksik oksijen türlerinin oluşmasına neden olmakta ve bu maddelerin zararlı etkileri ile karşı karşıya bırakmaktadır. Reaktif oksijen türleri (ROS) olarak adlandırılan bu toksik serbest radikaller, hücre için yaşamsal olan fizyolojik ve metabolik olaylar ile beraber oluşmaktadır (Gutteridge, 1984).
İnsülin hormonunun yokluğu veya yetersizliği sebebiyle açlık kan şekerinin normalin üzerinde seyrettiği, tüm yaş gruplarında görülebilen ve yaşam boyu süren, hiperglisemi ile birlikte özel komplikasyonlara yol açan diyabet, vücuttaki birçok organı etkileyen ve serbest oksijen radikallerinin patalojisinde yer aldığı bir hastalıktır. Vücuttaki glukoz miktarına bağlı olarak glukozun artan otooksidasyonu (Wolff ve Dean, 1987), enzimsel olmayan protein şekerlenmesi (Ceriello, 1997) ve polyol yolundaki enzim aktivitelerinin artması (Williamson ve ark., 1993) diyabetli dokularda serbest oksijen radikallerin artmasına neden olmakta, dolayısı ile hücrelerin enzimsel ve enzimsel olmayan antioksidan defans sistemlerini etkilemektedir (Brownlee, 2001).
Protein fosforilasyonu hücresel işlevlerin kontrolündeki belli başlı yollardan biri olarak bilinmektedir. Redoks düzenleme; hücresel yükseltgenme ve indirgenme potansiyel farklılıklarının değişmesi ile proteinlerin ve/veya enzimlerin aktivitelerinin değişmesi ve hücresel aktivitelerin kontrol edilmesi olarak tanımlanabilmektedir. Hücre içi redoks denge birçok mekanizma ile kontrol edilmekte olup, bu dengenin bozulması çeşitli enzimlerin gen ve protein ekspresyonlarında değişikliklere yol açabilmektedir (Schoonbroodt ve Piette, 2000).
Bu çalışmanın amacı, diyabet hastalığında varlığı ya da etkileri ispatlanmış olan oksidatif stresin karaciğer dokularında toplam protein fosforlanmasına etkilerini
araştırmak, meydana gelen değişikliklerin C vitamini (VC) ve resveratrol (RSV) gibi kuvvetli antioksidanlar ile nasıl değiştiğini ortaya çıkarmaktır. Bu çalışma ile aktivitelerinin fosforilasyon ile kontrol edildiği daha önceden bilinen katalaz (KAT) ve glutatyon peroksidaz (GPx) enzimlerinin fosforilasyon düzeylerinin diyabet ve antioksidan uygulaması ile nasıl değiştiği araştırılmış ve post-translasyonel bir mekanizma ile düzenlendiği savunulan bu enzimlerin düzenlenme mekanizmaları ortaya çıkarılmaya çalışılmıştır.
2. KURAMSAL TEMELLER ve KAYNAK ARAŞTIRMASI
2.1. Serbest Radikaller
Bir molekül ya da molekül gruplarının en dış yörüngesinde eşlenmemiş bir elektron taşıyorsa bu molekül ya da molekül gruplarına serbest radikal adı verilir. Molekülün bu radikal özelliği, kimyasal simgesinin sağ üst köşesine konulan nokta ile belirtilir. Serbest radikaller, mevcut paylaşılmamış elektronlarından dolayı oldukça reaktif atom ve moleküllerdir.
Oksijen; doğada dioksijen (O2) olarak bulunan sekiz atom numaralı kararsız bir
elementtir. Oksijenin bu kararsız durumu enerji düzeylerinde bulunan elektronların yapısına bağlıdır. Oksijen molekülünün son orbitali önemlidir. Bu orbitallerden herhangi birindeki elektron, bir orbitalden diğerine geçtiğinde veya farklı orbitallerde farklı yönlerde döndüğünde ‘singlet oksijen’ (ıO2) oluşur. Orbitalden birine ters dönen
bir elektron veya ikisine ters dönen iki elektron gelmesi sonucu ‘oksijen radikali’ meydana gelir. Oksijen molekülünde oluşan radikal, eşlenmemiş tek elektronundan dolayı çok dengesiz olup hızla ortamdan uzaklaşmak vasıtasıyla elektronlarını kolaylıkla bir başka moleküle verebilir (redüksiyon) veya bir elektron almasıyla elektron çifti oluşturabilir (oksidasyon). Bu durum sonucunda da radikal forma dönüşürler (Hochstein ve Atallah, 1988; Karabiga, 2006). Organizmalarda oluşabilen serbest oksijen türleri (ROS) Çizelge 2.1’de özetlenmiştir.
Çizelge 2.1. Serbest Oksijen Radikalleri (Karabiga, 2006)
1- Oksijen Merkezli Serbest Radikaller ▪ Moleküler Oksijen
Üçlü (triplet) durum 3
O2
Tekli (singlet) durum 1
O2
▪ Süperoksit Radikali O2 -·
▪ Hidroksil Radikali OH·
▪ Perhidroksi Radikali HO2·
▪ Alkoksi Radikali RO·
▪ Peroksi Radikali ROO·
2- Oksijen Merkezli Olmayan Serbest Radikaller ▪ Karbon Merkezli Lipit Radikalleri L· Alkil Radikalleri R· ▪ Sülfür Merkezli Sülfür Radikalleri R-S ▪ Hidrojen Merkezli Hidrojen Atomu +H ▪ Demir Merkezli Perferil Radikalleri Fe3+- O2 - Fe2+ ▪ Azot Merkezli
Nitrik Oksit NO·
Nitrojen Dioksit NO2·
3- Radikal Olmayan Reaktif Türler
▪ Ozon O3
▪ Hidrojen Peroksitler
Hidrojen Peroksit H2O2
Lipit Peroksitleri LOOH
▪ Hipoklorik asit HOCl
▪ Kloraminler R’RNCl
Pek çok fizyolojik durum sonucu oluşan serbest oksijen radikalleri, antioksidan savunma mekanizmaları ile etkisiz hale getirilirler. Antioksidan savunma sistemi ve serbest oksijen radikalleri üretimi arasındaki denge bozulduğunda, mevcut serbest oksijen radikalleri seviyesi artar. Serbest oksijen radikalleri stabil olmayan moleküller olmakla birlikte organik ve inorganik kimyasallarla hızlı bir şekilde reaksiyona girerler.
Serbest oksijen radikalleri üretimini artıran nedenler arasında hücrenin normal metabolizması sırasında gerçekleşen oksidasyon-redüksiyon reaksiyonları (örneğin: solunum, ksantin oksidaz, fenton reaksiyonu, moleküldeki bağların parçalanması sonucu farklı elektronların farklı atomlar üzerinde kalması) gösterilebilir. Ayrıca diyabet, ateroskleroz, karsinogenez, postiskemik reperfüzyon hasarı, enfeksiyonlar, enflamasyonların yanısıra yaşlanma ile ilgili hastalıklarda serbest radikallerin üretiminin artmış olduğu çeşitli çalışmalarda da gösterilmiştir. (Young ve Woodside, 2001; Yarıktaş ve ark., 2003; Cherubini ve ark., 2005; Halifeoğlu ve ark., 2005; Karabiga, 2006).
Serbest radikaller vücutta önemli moleküllere (hücrelerde proteinlere ve genetik materyale) zarar verirler. Bundan başka hücre zarının harabiyetine, geçirgenliğinin artmasına ve sonuçta hücrenin ölümüne kadar giden bir seri olaya yol açabilirler (Fang ve ark., 2002). Mitokondriyal, endoplazmik ve nükleer elektron taşıma sistemlerinde (sitokrom P-450), peroksizomlarda, monosit ve nötrofillerin fagositozu gibi normal metabolik olaylar sırasında çok miktarda serbest radikal üretilmektedir (Da Ros ve ark., 2004). Bu nedenle serbest radikalleri oluşturan yapı ve mekanizmalar endojen ve ekzojen olmak üzere ikiye ayırabiliriz (Çizelge 2.2).
2.1.1. Serbest Radikal Çeşitleri
Süperoksit (O2-.)
Süperoksit radikali, doğal oksijen molekülünün başka bir molekülden elektron almış hali olup oksijenden kaynaklanan tüm radikaller içinde en çok ve en kolay oluşanıdır. Süperoksit radikalinin bu özelliği, oksijenin suya indirgenmesi olayında meydana gelen ilk radikal olmasındandır (Halliwell, 2010).
Çizelge 2.2. Hücredeki serbest oksijen radikali kaynakları (Tekkes, 2006)
Endojen Kaynaklar Ekzojen Kaynaklar
Mitokondriyal elektron transport zinciri İlaç oksidasyonları (Ör. Parasetamol, CCl4)
Kloroplast elektron transport zinciri İyonize radyasyon
Oksidan enzimler: Güneş ışığı
Ksantin oksidaz X- ışınları İndolamin dioksijenaz UV- ışınları Triptofan dioksijenez Isı şoku
Galaktoz oksidaz Glutatyonu okside eden maddeler Siklooksijenaz Ortam havası:
Lipooksijenaz Sigara dumanı
Mono aminooksidaz Ozon
Fagositik hücreler: Kükürtdioksit
Nötrofiller Egzoz gazları
Monosit ve makrofajlar Eozinofiller
Endotelyal hücreler
Oto-oksidasyon reaksiyonları (Fe+2, epinefrin)
Negatif yüklü olması nedeniyle anyon kanallarını kullanarak ya da lipit tabakalarından difüze olarak plazma membranını geçebilen süperoksit, böylece uzak mesafelere ulaşıp hücresel elemanlara zarar verebilmektedir. Mitokondriyal elektron transfer zincirinde redükte nikotinamid adenin dinükleotid (NADH)’ın okside nikotinamid adenin dinükleotid (NAD+)’a dönüşürken de üretilebilmektedir (Şekil 2.1).
Şekil 2.1. Mitokondriyal elektron iletim zinciri tarafından süperoksitin üretimi (Brownlee,
2001)
Süperoksit radikali bunun dışında pek çok oksidaz tarafından ve ortamda bulunan geçiş metallerinin otooksidasyonu sonucunda da oluşabilmektedir (Eşitlik 2.1 ve 2.2).
Fe+2 + O2 Fe+3 + O2.- (2.1)
Cu+ + O2 Cu+2 + O2.- (2.2)
Ortamda artmış olan süperoksit düzeyleri, süperoksit dismutaz (SOD) enzimi ile hidrojen peroksit (H2O2) ve oksijene dönüştürülerek yok edilir (Eşitlik 2.3).
SOD
2O2.- + 2H+ H2O2 + O2 (2.3)
Süperoksit dismutaz enzimi ile hücresel süperoksit düzeyleri sıkı kontrol altına alınmaktadır (Cherubini ve ark., 2005; Tekkes, 2006).
Hidrojen Peroksit (H2O2)
Normal oksijen molekülü başka herhangi bir molekülden iki elektron almışsa peroksit adı verilen molekül oluşmaktadır. Eğer peroksit molekülü, iki H molekülü ile birleşirse hidrojen peroksit (H2O2) adı verilen ve diğer önemli ve çok aktif radikalleri
oluşturabilen yapı ortaya çıkmaktadır (Şekil 2.2).
Şekil 2.2. Hidrojen peroksitin moleküler şekli
H2O2,SOD enzimi ile veya Eşitlik 2.4’te olduğu gibi spontan olarakta oluşabilmektedir.
2O2. + 2H+ H2O2 + O2 (2.4)
Normalde H2O2 bir radikal değildir. Ancak bu molekül, membranları geçen,
sitoplazmaya diffüze olan ve uzun süre kalabilen bir oksidan olarak bilinir. Bu özelliğinden dolayı süperoksitin geçemediği ve membranla korunan yapılara kolaylıkla ulaşabilir. Diffüze olup geçtiği dokularda en reaktif ve zarar verici radikal olan hidroksil radikali oluşturmak üzere (Eşitlik 2.5) kolaylıkla yıkılabilir ve bunu da süperoksitle reaksiyona girerek yapar.
H2O2 + O2.- OH. + OH- + O2 (2.5)
Ayrıca hidrojen peroksit, serbest halde bulunan Fe+2
ile reaksiyona girmesi sonucu hidroksil radikaline dönüşür (Eşitlik 2.6 ve 2.7) (Vincent ve ark., 2004; Cherubini ve ark., 2005).
O2.- + Fe+3 O2 + Fe+2 (2.6)
Ayrıca, aminoasit oksidaz, ksantin oksidaz (XO) gibi bazı oksidaz enzimlerinin faaliyeti sonucunda da in vivo olarak H2O2 üretilir (Şekil 2.3) (Selen İşbilir, 2008).
Şekil 2.3. Ksantin oksidaz (XO) ile H2O2 üretimi
Hidroksil Radikali (OH.)
Bilinen en zarar verici radikal olan hidroksil radikali, bir elektronu eksik olan tek atomlu oksijen ile H+ ’in birleşmesinden oluşur. Oksijen radikalleri içinde en reaktif olanı olmakla beraber, yarı ömrü en kısa olan radikaldir. Bu özelliğinden dolayı kaynağından fazla uzaklaşmayarak çevresinde bulunan en yakın hedefleri etkilerler. Hidroksil radikali, hidrojen peroksitin geçiş metalleriyle indirgenmesi sonucu (Eşitlik 2.9) veya süperoksit radikaliyle reaksiyona giren hidrojen peroksit ile (Eşitlik 2.10) oluşabilmektedir. Bu reaksiyon katalizörsüz çok yavaş olmasına rağmen Fe+2
katalizörlüğü ile çok hızlı gerçekleşmektedir.
O2.- + Fe+3 O2 + Fe+2 (2.8)
Fe+2 + H2O2 Fe+3 + OH. + OH- (Fenton reaksiyonu) (2.9)
H2O2 + O2.- OH. + OH- + O2 (Haber-Weiss reaksiyonu) (2.10)
Hidroksil radikali, bir çok yapıyla örneğin, aminoasitler, nükleik ve organik asitler, fosfolipitler ve şekerler gibi biyokimyasal maddelerle reaksiyona girerek bu moleküllerin yapısını bozabilmektedir (Gönenç, 1995; Young ve Woodside, 2001; Vincent ve ark., 2004; Cherubini ve ark., 2005; Tekkes, 2006).
Nitrik Oksit (NO.)
Nitrik oksit, hem hücre içi hem de hücre dışındanda düzenleyici işlev gören küçük, reaktif bir serbest radikal moleküldür. Nitrik oksitin moleküler orbitalinde eşlenmemiş elektron çifti bulunduğundan dolayı bir serbest radikal olarak yarı ömrü çok kısadır. NO. oluşumu memeli hücrelerinde yaygın olarak NADPH’a bağımlı sitozolik bir enzim olan nitrik oksit sentaz (NOS) enziminin aktivitesi ile oluşur (Benzer ve Temüzer Ozan, 2003; Vincent ve ark., 2004; Pratic`o, 2005). Birçok hücresel işlem; vasküler düz kas hücreleri, endotel, trombosit, nöronlar ve diğer NO üreten hücrelerden salınan NO tarafından düzenlenmektedir (Bülbül ve Soylu, 2008).
2.1.2. Serbest Radikallerin Etkileri
Güçlü reaktif özelliklere sahip olan serbest radikaller, hücresel bütün yapıları; DNA, protein, lipit, karbonhidrat ve enzim sistemleri gibi tüm önemli bileşikleri kolayca etkileyerek yıkıma uğratabilirler (Selen İşbilir, 2008). Çok reaktif olan hidroksil radikali ve süperoksit radikali sitoplâzma, nükleus, mitokondri ve endoplazmik retikulum membranlarında lipit peroksidasyonuna sebep olabilmektedir (Şekil 2.4) (Gürbüz, 2008).
Serbest Radikallerin Proteinler Üzerine Etkileri
Proteinler, büyük molekül ağırlığına sahip polipeptitler olup hücre foksiyonlarında ve hücre yapısında çok önemli görevler üstlenmekle birlikte, oksidatif reaksiyonlar sonucu önemli modifikasyonlara uğrayabilmektedir. Proteinlerin yapıtaşı olan aminoasitler; serbest radikallerin en önemli hedeflerindendir. Proteinleri oluşturan aminoasitlerin hasar görmesi, proteinlerde kalıcı değişikliklere sebep olur. Sistein, sistin, histidin, metiyonin, tirozin ve triptofan aminoasitleri proteinler içerisinde serbest radikallere en duyarlı moleküllerdir ve serbest radikaller ile tepkimeleri sonucu Şekil 2.5’te belirtilen yan ürünlere dönüşebilmektedirler.
2-Pirolidon Glutamik semialdehit Aminokadipik semialdehit
2-Amino-3-ketobutirik asit
Şekil 2.5. Serbest radikaller tarafından oluşabilen proteinler üzerindeki karbonilasyon ürünleri
Proteinlerin serbest radikaller ile oksidasyonu sonucu hasarına karşı duyarlılığı, aminoasit bileşimine, protein aktivasyonundan veya yapısal düzenlenmesinden sorumlu aminoasitlerin dizilimine ve hasarlı proteinlerin onarılabilirliğine bağlıdır (Karabiga, 2006). Serbest radikallerin proteinler üzerinde yol açtığı başlıca değişiklikler aminoasitlerin modifikasyona uğraması, proteinlerin fragmantasyonu ve çapraz bağların oluşması şeklinde sıralanabilir (Kayış, 2010).
Serbest Radikallerin Karbonhidratlar Üzerine Etkileri
Serbest radikaller, karbonhidratlar üzerinde de ciddi etkiler bırakmaktadır. Fizyolojik şartlarda otooksidasyona uğrayan glikoz, mannoz ve deoksi şekerler, süperoksit ve hidrojen peroksitin oluşumuna sebebiyet vermektedir. Monosakkaritlerin otooksidasyonu, protein çapraz bağlanmalarına yol açarak bazal membranda kalınlaşmaya yol açarak katarakt ve benzeri hastalıklara sebep olabilmektedir (Tekkes, 2006).
Reaktif oksijen türlerinin karbonhidratlar üzerinde polisakkarit depolimerizasyonunun yanında özellikle monosakkarit otooksidasyonu gibi önemli etkileri de bulunmaktadır. Monosakkaritlerin otooksidasyonu sonucu oluşan hidrojen peroksit, süperoksit ve okzoaldehitler diyabet ve sigara içimi ile ilgili patolojik olaylarda rol oynar (Selen İşbilir, 2008).
Serbest Radikallerin Nükleik Asitler Üzerine Etkileri
İyonize edici radyasyona bağlı hücre ölümünün temel nedeni nükleik asitlerin ROS ile reaksiyona girmesidir. ROS nükleik asit baz değişimlerine veya DNA çift sarmalının hasar görmesine, bu da kromozomal mutasyonlara ve sitotoksisiteye neden olabilmektedir (Ames ve ark., 1993; Frei, 1994). Ayrıca aktivite gösteren nötrofillerden kaynaklanan hidrojen peroksit, membrandan kolayca geçerek hücre çekirdeğinde nükleik asit hasarlarına sebep olmaktadır (Ames ve ark., 1993; Cheesman ve Slater, 1993).
En tehlikeli serbest radikal çeşitlerinden biri olan hidroksil radikali, deoksiriboz ve bazlarla kolayca reaksiyona girerek bu moleküllerin yapılarını değiştirebilmektedir. Hidroksil radikali eğer DNA’nın yakınında oluşursa pürin ve primidin bazlarına saldırarak baz değişikliklerine ve mutasyonlara neden olabilmektedir (Şekil 2.6).
Şekil 2.6. Hidroksil radikalinin pürin ve pirimidin bazları üzerine etkisiyle oluşan son ürünler
(Selen İşbilir, 2008)
Aynı zamanda hidroksil radikali ile karşılaştırıldığında nükleik asitlerle reaksiyon oluşturma yeteneği daha az olan singlet oksijen, guanin gibi yüksek elektron yoğunluklu bölgeler içeren moleküllerle tepkimeye girerek nükleik asitlerin yapısını bozabilmektedir (Kayış, 2010).
Serbest Radikallerin Lipitler Üzerine Etkileri
Bütün biyomoleküller serbest radikallerin hedefindedir ve ciddi bir şekilde etkilenirler. Serbest radikallerin yıkıcı etkilerinden en çok etkilenen biyomoleküller olan lipitlerin hücre içerisinde en çok bulunduğu bölgeler hücre zarlarıdır. Bu nedenle hücre zarları serbest radikal kaynaklı oksidasyonlara çok duyarlı biyomoleküllerdir (Eşitlik 2.11) (Cheesman ve Slater, 1993; Tekkes, 2006). Atmosferik oksijen varlığında serbest radikaller lipitler üzerinde lipit peroksidasyonu tepkimelerini başlatarak peroksi lipitleri oluşturmaktadır (Eşitlik 2.12-2.14).
LH + R. L. + RH (2.11) L. + O2 LOO. (Lipit peroksit radikali) (2.12)
LOO. + LH LOOH + L. (2.13)
LOOH LOO. ; LO. (2.14)
Lipit peroksidasyonu süperoksit radikali, hidroksil radikali, peroksil radikali ve alkoksil radikali gibi kuvvetli radikaller ile başlamaktadır. Bu radikaller çeşitli mekanizmalarla hücrede oluşturulmakta ve hücre membranının harabiyetine yani lipit peroksidasyon ürünlerinin oluşmasına neden olabilmektedir (Şekil 2.7). Bununla beraber demir iyonunun da lipit peroksidasyonunda önemli rolü bulunmaktadır (Karabiga, 2006).
Şekil 2.7. Serbest oksijen radikallerin oluşumu ve lipitlere etkileri
Serbest radikaller hücre membranında bulunan yağ asitleri ve kolesterolün doymamış bağları ile reaksiyona girip peroksidasyon reaksiyonlarını başlatabilmektedir. Burada ilk önce yağ asidi kendi üzerinde bir elektron kalacak şekilde parçalanmakta ve lipit radikalini oluşturmaktadır. Oluşan lipit radikali atmosferik oksijen ile reaksiyona girerek lipit peroksit radikalini oluşturur ve böylece lipit peroksit radikali de ortamdaki diğer doymamış yağ asitleriyle reaksiyona girerek zincirleme bir reaksiyon oluştur.
Ayrıca lipit peroksitleri ortamda bulunun ve yağ asitlerinin parçalanmasıyla oluşmuş olan üzerinde bir elektron bulunan hidrojen atomları ile reaksiyona girerek lipit hidroperoksidlerini de oluşturabilmektedir (Şekil 2.8) (Young ve Woodside, 2001; Niki ve ark., 2005).
Şekil 2.8. Lipit peroksidasyonunun kimyasal mekanizması (Tekkes, 2006)
Çok Doymamış Yağ Asidi
Lipit Radikali
Dien Konjugat
Lipit Peroksil Radikali
Endoperoksit Hidroperoksit
Oluşan lipit peroksitleri daha sonra yıkım ürünleri olan malondialdehit (MDA) ve 4-hidroksi nonenal gibi yapılara dönüşerek DNA ve/veya proteinlerle reaksiyona girerek DNA hasarı ve protein bozukluklarına yol açabilir. Lipit peroksidasyonu sonucunda membranlarda yapısal ve fonksiyonel hücre hasarı oluşmaktadır (Young ve Woodside, 2001; Masella ve ark., 2005; Niki ve ark., 2005). Yapılan çalışmalarda diyabet, kanser, alzaymır gibi bir çok hastalıkta plazma ve doku lipit peroksidasyon ürünlerinde artış meydana geldiği bulunmuştur (Halifeoğlu ve ark., 2005).
2.2. Vücudun Antioksidan Savunma Mekanizmaları
Reaktif oksijen türlerinin vücutta meydana getirdiği hasarları ortadan kaldırmak için vücutta görev alan savunma sistemlerine antioksidan savunma sistemleri adı verilir. Antioksidanlar, hem doğrudan hem de dolaylı olarak ilaçların, karsinojenlerin ve bir çok toksik radikal reaksiyonlarının istenmeyen etkilerine karşı hücreleri koruyan savunma sistemleridir (Mercan, 2004; Halifeoğlu ve ark., 2005).
Antioksidanlar, enzimatik olan (antioksidan enzimler) ve enzimatik olmayan (antioksidan bileşikler) antioksidanlar olmak üzere iki ana gruba ayrılırlar (Çizelge 2.3). Antioksidan enzimler; katalaz (KAT), süperoksit dismutaz (SOD), glutatyon peroksidaz (GPx), ve glutatyon redüktaz (GR) gibi enzimlerden oluşmaktadır. Antioksidant bileşikler ise Vitamin A, C, E, poliaminler, melatonin, NADPH, adenozin, bilirubin, koenzim Q-10, ürat, ubikinon, metalotionin, polifenoller, flavonoidler, fitoöstrojenler, sistein, homosistein, taurin, metiyonin, s-adenozil-L-metiyonin, resveratrol, nitroksidler, karotenoidler (beta-karoten gibi), ksantofiller, melatonin, seruloplazmin, hemoglobin, glutatyon ve tokoferoller gibi çok sayıda bileşikten oluşmaktadır (Granado ve ark., 1998; Jemal ve ark., 1998; Akgül ve ark., 1999; Yarıktaş ve ark., 2003; Mercan, 2004; Halifeoğlu ve ark., 2005).
Çizelge 2.3. Antioksidan Savunma sistemleri (Karabiga, 2006)
ANTİOKSİDAN SAVUNMA SİSTEMLERİ
Enzim Sistemi Radikal Tutucular
Süperoksit Dismutaz Suda Çözünenler
Glutatyon Peroksidaz Glutatyon
Glutatyon Redüktaz Vitamin C
Katalaz Sistein
Glukoz 6-P-Dehidrogenaz Ürik asit
Glutatyon-S-Transferaz Resveratrol Yağda çözünenler Vitamin E Beta-karoten Bilurubin Flavonoidler
Metal iyonu Bağlayanlar Transferin Seruloplazmin Haptoglobülin Albumin Hemopeksin Ferritin
Serbest radikaller ve olası hasarları, antioksidan bileşiklerle beraber antioksidan enzimler tarafından denetlenmektedir. Organizmada meydana gelen serbest radikalleri sürekli olarak zararsız hale getirmeye çalışan süperoksit dismutaz (SOD), katalaz (KAT), glutatyon peroksidaz (GPx), glutatyon S-transferaz (GSTs) gibi çeşitli enzimler bulunmaktadır (Şekeroğlu, 2000; Çiğremiş ve ark., 2003).
SOD enzimi hem sitoplazmada hem de mitokondride süperoksit radikallerini hidrojen peroksite çevirmektedir. Peroksizomal KAT ve sitoplazmik GPx enzimleri, oluşan hidrojen peroksiti hidroksi radikaline dönüşmeden nötralize etmektedirler (Şekil 2.9).
Şekil 2.9. Serbest radikallerin oluşumu ve antioksidan enzimlerin etkisi (Vincent ve ark., 2004)
GPx enzimi antioksidan molekül olan indirge glutatyonu, hidrojen peroksiti indirgemede kullanmaktadır. Yine aynı şekilde sitoplazmik GST enzimleri indirge glutatyonu kullanarak hücrelerdeki oksidan stresi azaltmaya yönelik görev almaktadırlar. GST’lar aynı zamanda oksidatif olarak modifiye edilmiş molekülleri hücre içerisinde detoksifiye ederek, hücresel peroksitleri etkisiz hale getirirler (Şekil 2.10) (Kwag ve ark., 1999).
Şekil 2.10. Antioksidan enzimlerin glutatyon redoks döngüsü ile beraber serbest radikal
2.2.1. Enzimatik Antioksidanlar
Süperoksit Dismutaz (SOD) (EC: 1.15.1.1)
İlk defa McCord ve Fridovich tarafından 1969 yılında süperoksit radikallerini tepkimeye sokarak hidrojen peroksit ve moleküler oksijene dönüşümünü sağlayan ve lipit peroksidasyonunu inhibe eden bir metaloenzim olarak tanımlanmıştır (Moscone, 1988). Bu reaksiyon kendiliğinden gerçekleşebildiği gibi, SOD eşliğinde (Eşitlik 2.15) 4000 kat daha hızlı oluşmaktadır (Kayış, 2010).
SOD
O2.- + O2.- + 2H+ H2O2 + O2 (2.15)
Normal metabolizma sırasında hücrelerde yüksek oranda süperoksit radikali oluşmasına rağmen bu oran SOD sayesinde çok düşük seviyelerde tutulmaktadır. Reaksiyon sonunda membrandan geçemeyen süperoksit radikali membranlardan geçebilme özelliğine sahip hidrojen peroksite dönüşerek geçiş metalleri varlığında Fenton ve Haber-Weiss reaksiyonları ile en tehlikeli radikal olarak bilinen hidroksil radikallerine dönüşmektedir (Rigo ve ark., 1977). Bu nedenle SOD aktivitesindeki artış ile ortamda aşırı H2O2 oluşmasının ancak KAT ve GPx enzimlerinin artan aktiviteleri ile kontrol
edilebileceği gösterilmiştir (Kayış, 2010).
SOD’un aktif merkezinde bulunan geçiş metallerinin farklı olması sonucu, Cu/Zn-SOD, Mn-SOD ve Fe-SOD olmak üzere üç çeşide ayrılır. Cu/Zn-SOD hücrelerde en bol bulunan SOD izomeri olup toplam 32 kDa molekül ağırlığına sahip iki eşit molekül ağırlıklı alt üniteden oluşmakta ve her alt ünitesinde de bir bakır ve bir çinko atomu içermektedir (Mruk ve ark., 2002). Cu/Zn-SOD hayvan hücrelerinde en çok sitozolde bulunmakla beraber lizozomlarda, iç ve dış mitokondrial membran boşluklarında ve çekirdekte bulunduğu tespit edilmiştir.
Cu/Zn-SOD’lar için siyanür iyonları güçlü bir inhibitördür, aynı zamanda dietilditiyokarbamatlar enzimin aktif bölgesinde bulunan bakır iyonunu bağlayarak aktif bölgeden uzaklaştırmak suretiyle enzimi çalışamaz hale getirmektedir (Kayış, 2010). Mn-SOD hayvan bakteri ve bitki dokularında bulunabilmekle beraber mitokondriyal bir enzim olup eşit molekül ağırlıklı dört alt üniteden oluşmaktadır. Aktif bölgesinde
süperoksit radikalinin aynı organelde ortadan kaldırılmasından sorumludur (Orbea ve ark., 2000).
Fe-SOD, 41 kDa ağırlığında iki alt üniteden oluşan ve her alt ünitesinde bir ya da iki demir atomu bulunduran bir enzimdir. Hücre dışı bir enzim olan Fe-SOD, ekstrasellular matriks içerisinde süperoksit radikaline karşı savaşmaktadır (Kayış, 2010).
Katalaz (KAT) (EC: 1.11.1.6)
Katalaz (KAT) enzimi glutatyon peroksidaz (GPx) ile beraber hücre içi hidrojen peroksiti detoksifiye eden, yapısında prostetik grup olarak porfirin halkası ve dolayısıyla demir içeren peroksizomal bir enzimdir. KAT enziminin temel fonksiyonu, serbest radikallerin oluşturabilecekleri geri dönüşümsüz hasarların önüne geçmek için moleküler oksijen varlığında hücre metabolizmasının bazı basamaklarında sentezlenen, hidrojen peroksit veya ROOH gibi peroksitlerin radikal oluşturmadan ortadan kalkmalarını sağlamaktır. Eşitlik 2.16’da gösterildiği gibi KAT, iki peroksit molekülünü su ve moleküler oksijene dönüştüren hücresel antioksidan enzimlerin en önemlilerinden bir tanesidir.
KAT
H2O2 + H2O2 2H2O + O2 (2.16)
Hücre içerisinde sitozolde de varlığı tespit edilmekle birlikte en çok peroksizomlarda bulunmaktadır. KAT daha çok peroksizomlarda bulunurken, GPx ise sitozol ve mitokondride lokalize olmasıyla birbirlerini tamamlayıcı bir yerleşim gösterirler. Böylece hücre içi hidrojen peroksitin konsantrasyonunu tamamlayıcı birliktelik çerçevesinde etkin bir şekilde düzenlerler.
Yapılan çalışmalar birçok hastalıkta örneğin Down sendromu, kanser ve anemide serumda KAT aktivitesinin değiştiğini göstermektedir (Benzer ve Temüzer Ozan, 2003).
Glutatyon Peroksidaz (GPx) (EC: 1.11.1.9)
Glutatyon peroksidaz enzimi ilk kez 1957 yılında Mills tarafından hayvan eritrositlerinden izole edilmiş, mitokondri, sitozol ve hücre membranlarında bulunan ve glutatyonu elektron kaynağı olarak kullanarak hidrojen peroksit ve organik hiperoksitlerin indirgenmesinden sorumlu bir antioksidan enzimdir (Eşitlik 2.17 ve 2.18).
GPX
2GSH + H2O2 GSSG + 2H2O (2.17)
GPX
2GSH + ROOH GSSG + ROH + H2O (2.18)
Normal koşullarda KAT ve GPx, hücrelerin farklı yerlerine yerleştiklerinden dolayı, karaciğerde endojen olarak oluşan H2O2 seviyesini düzenlemede birlikte çalıştıkları
bilinmektedir (Selen İşbilir, 2008).
GPx enzimi aktif bölgesinde selenosistein formunda, kovalent bağlı selenyum atomu içerir. GPx organik hidroperoksitler ile birlikte aynı zamanda H2O2’ye karşı aktiftir.
GPx H2O2’de dahil çeşitli hidroperoksitlerin yıkımını GSH’ın oksidasyonu ile
gerçekleştirir. Bu reaksiyon ile oluşan okside glutatyon (GSSG) ise glutatyon redüktaz (GR) enzimi yardımı ile tekrar eski yapısına, yani GSH’a indirgenir (Kayış, 2010). Fosfolipit savunmasını yapan selenyuma bağımlı GPx’in bir diğeri ise fosfolipit hidroperoksit GPx (PL-GPx) formudur. Bu enzim bir selenyum atomu içerir ve fosfolipit hidroperoksitleri alkollere indirgeyerek membran koruyucu özelliğindeki en önemli antioksidan bileşik olan E vitamini eksikliğinde membranı radikallere karşı korumaktadır (Kayış, 2010).
Glutatyon Redüktaz (GR) (EC: 1.8.1.7)
Glutatyon redüktaz, bir flavin enzimidir. Koenzimi NADPH ve prostetik grubu FAD’tir. GR sitozol ve mitokondride görev yapmaktadır (Karabiga, 2006). GPx vasıtasıyla hidroperoksitlerin indirgenmesi ile oluşan okside glutatyonun (GSSG) tekrar indirgenmiş
glutatyona (GSH) dönüşümünü katalizler. Bu reaksyonun oluşabilmesi için NADPH gereklidir (Şekil 2.11) (Gürbüz, 2008).
Şekil 2.11. Antioksidan enzimler ve glutatyonun indirgenme reaksiyonu (Gürbüz, 2008)
Glutatyon-S-Transferaz (GST) (EC: 2.5.1.18)
Glutatyon-S-Transferaz (GST), her biri iki protein alt birimden oluşmuş bir enzim ailesidir. Genel olarak bir mikrozomal ve üçü de sitozolik olmak üzere 4 ana gruba ayrılırlar. GST katalitik ve katalitik olmayan çok sayıda fonksiyonda işlevi bulunmaktadır. Çeşitli şekillerde organizmaya giren ksenobiyotiklerin biyotransformasyonunda önemli roller üstlenmektedir. Başta araşidonik asit ve linoleat hidroksiperoksitleri olmak üzere lipit hidroperoksitlere karşı GST’ler Se-bağımsız GPx aktivitesi göstererek bir antioksidan savunması oluştururlar (Eşitlik 2.17) (Selen İşbilir, 2008; Storey, 1996).
GST
2.2.2. Enzimatik Olmayan Antioksidanlar
Glutatyon (GSH)
Glutatyon (Şekil 2.12), serbest radikaller ve peroksitlerle reaksiyona girerek hücreleri oksidatif hasara karşı koruyan antioksidan bir bileşiktir (Reiter, 1997).
Şekil 2.12. Glutatyonun molekül şekli
Serbest radikallere karşı önemli bir antioksidan olan glutatyon, glutamik asit, sistein ve glisinden meydana gelen bir tripeptiddir (Kayış, 2010). GSH hücrede sitozol, mitokondri ve nükleusta fazla miktarda bulunmakla beraber hücre içinde büyük bir kısmı indirgenmiş olarak (tiyol) bulunurken, daha az bir kısmı ise okside glutatyon (GSSG) şeklinde bulunur (Mytilineou ve ark., 2002).
GSH’a antioksidan özelliği sağlayan kısım tiyol grubudur. Glutatyon hücreyi oksidatif hasarlara karşı korumak için hidroksil ve singlet oksijen gibi reaktif oksijen türlerini temizlemenin yanında diğer serbest radikaller ve peroksitlerle de reaksiyona girebilmektedir (Kayış, 2010).
Bunun dışında hemoglobinin oksitlenerek methemoglobine dönüşümünün engellenmesine katkı yapar. Ayrıca fonksiyonel proteinlerin ve enzimlerin inaktivasyonunu engellemek için proteinlerdeki sülfidril (-SH) gruplarını redükte halde tutmanın yanında bu grupları oksidasyona karşı korur. Glutatyon, yabancı bileşiklerin detoksifikasyonu ve aminoasitlerin hücre içine taşınmasının yanında çeşitli metabolik fonksiyonlarda da rol oynar. Ayrıca oksidatif strese karşı eritrositleri, lökositleri ve göz lensini korumada hayati derecede önemli bir yer tutmaktadır (Reiter, 1997). GSH, protein konformasyonu, membran transportunun düzenlenmesi ve enzim aktivitesinin ayarlanması gibi mekanizmalarda da görev almaktadır (Kayış, 2010).
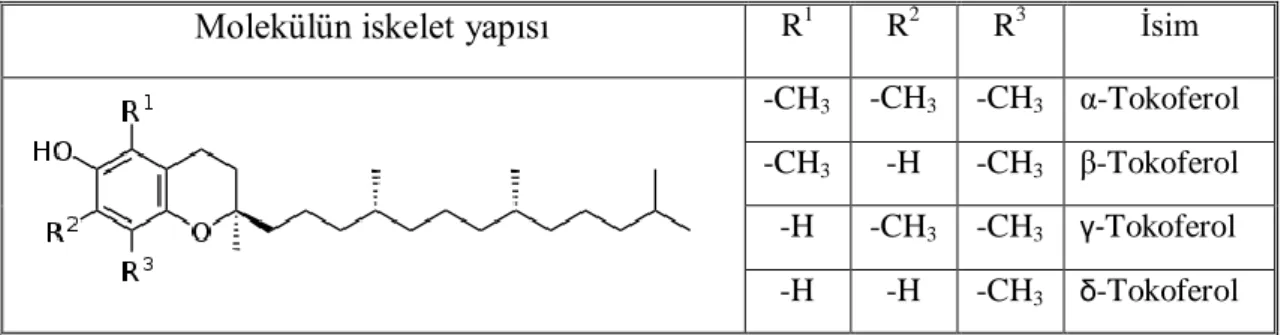
E vitamini (Tokoferol)
E vitamini, α, β, γ, δ-tokoferol olarak adlandırılan bileşiklere verilen genel bir isimdir. E vitamini oluşturan α, β, γ, δ-tokoferol bileşikleri arasında doğal dağılım olarak en çok bulunan ve biyolojik aktivite yönündende en yüksek aktiviteye sahip olan α-tokoferoldür (Şekil 2.13).
Molekülün iskelet yapısı R1 R2 R3 İsim
-CH3 -CH3 -CH3 α-Tokoferol
-CH3 -H -CH3 β-Tokoferol
-H -CH3 -CH3 γ-Tokoferol
-H -H -CH3 δ-Tokoferol
Şekil 2.13. E vitamininin değişik formlarının molekül şekli
α-Tokoferol, hidroksil, süperoksit, singlet oksijen radikalleri ile beraber lipit peroksil ve diğer bazı serbest radikallerin temizlenmesinde görev alır (Chow, 1991; Blokhina ve ark., 2003). Lipit peroksidasyonuna karşı ilk savunma hattını oluşturarak membranda serbest radikal toplayıcı etki oluşturuyor ve hücre membranında bulunan doymamış yağ asitlerini serbest radikallerin zararlı etkilerinden koruyarak, lipit peroksidasyonunu erken safhalarında engeller.
Ayrıca α-tokoferol, GPx ile birlikte çalışarak birbirlerini tamamlayıcı antioksidan etki gösterirler. Burada α-tokoferol peroksitlerin oluşumunu engellerken, GPx oluşmuş olan peroksitleri süpürür (Kayış, 2010).
C vitamini (Askorbik asit)
Aynı zamanda askorbat yada askorbik asit olarak da bilenen C vitamini (VC), kapalı formülü C6H8O6 olan bir bileşik olup organizmada birçok hidroksilasyon reaksiyonunda
indirgeyici güç olarak görev alır (Kayış, 2010).
VC insanda sentezlenemeyen, suda çözünen bir yapıya sahip olup, güçlü indirgeyici aktivitesinden dolayı aynı zamanda kuvvetli bir antioksidandır (Şekil 2.14).
Şekil 2.14: VC’nin molekül şekli
Askorbik asit etkili olarak singlet oksijenin yanında H2O2, süperoksit, hidroksil,
hipoklorit ve peroksil radikallerini tutma özelliğine sahiptir. Sıvı ortamdaki tüm peroksil radikallerini plazma lipitlerine difüzyon ile geçmeden tutar ve bu şekilde lipit peroksidasyonunun başlamasını engellemiş olur. Membranlarda bulunan α-tokoferol ile reaksiyona girerek α-tokoferolün yenilenmesini sağlar (Özer ve Gönül, 2006; Selen İşbilir, 2008; Halliwell, 1994). VC, nötrofillerin sebep olduğu peroksidasyona karşı plazma lipitlerini korur ve güçlü bir hipoklorat gidericisi olması ile fagositoz için de önemli olduğu gösterilmiştir.
VC yüksek konsantrasyonlarda antioksidan aktivitesinin yanında, düşük konsantrasyonlarda prooksidan olarak da aktivite gösterdiği, ancak antioksidan özelliği prooksidan özelliğinden daha baskın olduğu bildirilmiştir (Halliwell, 1994). Bu özelliğiyle ferri demiri ferro demire indirgeyerek fenton reaksiyonunda H2O2 ile
etkileşmeye eğilimli olan ferro demir oluşturmakta ve reaksiyon sonucunda süperoksit radikalinin oluşmasına sebebiyet vermektedir (Bendich ve ark., 1986).
A vitamini
A vitaminin metabolik öncül maddesi olan β-karoten çok güçlü bir singlet oksijen temizleyicisi olarak bilinir (Şekil 2.15). Bununla beraber hidroksil, peroksil ve alkoksil radikalleri ile doğrudan reaksiyona girerek lipit peroksidasyon zincir reaksiyonunda engelleyici rol oynar. β-karoten düşük oksijen seviyelerinde etkili olmakla birlikte aynı zamanda daha yüksek oksijen seviyelerinde etkili olan E vitaminin antioksidan etkisinin tamamlayıcısı olarak çalışır (Kayış, 2010).
Şekil 2.15: A vitamininin molekül şekli
Resveratrol
Resveratrol (RSV) (3,4,5 trihidroksi sitilben), bitkilerde biyolojik olarak oldukça aktif olan fitoaleksin yapıda bir maddedir (Şekil 2.16). İlk olarak şaraplık üzümlerde (Vitis vinifera) tespit edilen RSV’nin dokuları mantar enfeksiyonlarından ve/veya ultraviyole ışınlardan koruduğu bulunmuştur. RSV’nin en çok bulunduğu bitki Polygonum cuspidatum olarak bilinen bitkilerin kökleridir. RSV yenilebilir bitkilerde nadir olarak bulunmakla beraber itadori çayı, yer fıstığı, fıstık, üzüm ve şarap ile besinlerden alınır.
Şekil 2.16: Trans-Resveratrol’ün molekül şekli
RSV’nin başlıca biyolojik etkileri, serbest radikalleri berteraf etme, antikanser, trombosit agregasyonunun inhibisyonu, vazorelaksiyon kapasitesinde artış, lipit metabolizmasının modülasyonu ve peroksidasyonun inhibisyonu, östrojenik aktivite, eikozanoit sentezi modülasyonu olarak sıralanabilir (Frémont, 2000).
Birçok araştırmada RSV’nin süperoksit ve hidroksil radikallerini yakalama özelliğinin olduğu gösterilmiştir. Ayrıca RSV biyolojik sistemlerde bulunan antioksidanların hücre içi konsantrasyonlarının sürdürülmesini de sağlamaktadır (De La Lastra ve Villegas, 2007).
Yapılan bir çalışmada RSV’nin insan lenfositlerinde glutatyon miktarını arttırdığı belirtilmiştir. Bu çalışmada insan lenfositlerinde RSV’nin GR, GPx ve GST gibi
glutatyon metabolizması ile ilgili enzimlerin aktivitelerini değiştirdiği gösterilmiştir (Das ve Maulik, 2006).
RSV mitogen ile aktive olan protein kinaz (MAPK) kaskadını harekete geçirebilme özelliğine sahiptir. RSV protein kinazların farklı düzeylerinde rol oynayabilmektedir. Protein kinaz C (PKC) fosforilasyonunu inhibe ederek biyokimyasal reaksiyonlara yön verme özelliğine sahip olan RSV, MAPK kaskadının etkinliğini sağlayan diğer kinazları, fosfoinozitol 3-kinaz (PI3K) fosforilasyonunu ve bunun yanında protein kinaz B’nin (Akt/PKB) fosforilasyonunu da inhibe etme yeteneğine sahiptir. RSV, fosforilasyonun yanında aynı zamanda bazı genlerin ifadelenmesini de azaltma özelliğine sahiptir. Ayrıca p65’in transaktivasyonunu ve fosforilasyonunu IKK, PKC gibi kinazları inhibe ederek baskılamaktadır (Delmas ve ark., 2005).
2.3. Oksidatif Stres
Oksidatif stres; antioksidan maddelerin azalmasıyla reaktif oksijen türlerinin (ROS) aşırı üretimi olarak veya serbest radikaller ve antioksidanlar arasındaki dengenin serbest radikaller lehine bozulması olarak tanımlanmaktadır. Serbest radikaller ile antioksidan savunma sistemi arasındaki hassas dengenin, peroksidan ve oksidan maddelerin lehine kayması oksidan stresin gelişmesine yol açmaktadır. Oksidatif stresin, doku hasarına yol açtığı, kanser, diyabet ve ateroskleroz gibi patolojik durumların gelişmesinde etkin olduğu gösterilmiştir (Reuter ve ark., 2010; Valko ve ark., 2006; Joshi ve ark., 2005; Chevion ve ark.,2000).
Yapılan araştırmalarda bu durumun ayrıca diyabetin makro ve mikrovasküler komplikasyonlara neden olduğu pek çok araştırıcı tarafından vurgulanmaktadır. Oksidatif stresin ayrıca glukoz metabolizması üzerinde çeşitli etkileri bulunmaktadır (Memisogullari ve ark., 2003; Vincent ve ark., 2004; Memişoğulları ve Bakan, 2004; Cherubini ve ark., 2005).
Oksidatif stresin artışı, serbest radikal üretimindeki artıştan kaynaklanabilmektedir (Noyan ve ark., 2004). Oluşan serbest radikallerin yıkıcı etkileri organizmada bulunan ve antioksidan savunma sistemi olarak bilinen bir sistem tarafından denetlenmektedir (Şekil 2.17). Organizmadaki oksidatif streste görülen herhangi bir artışın, antioksidan sa-vunma sistemindeki bir azalmadan ya da serbest radikal üretimindeki bir artmadan
Şekil 2.17. Reaktif oksijen türevleri ve antioksidan mekanizmaları (Karabiga, 2006)
2.4. Diyabet
Diyabet, insülin salgılanmasında veya dokunun insüline yanıt vermesinde oluşan bozukluklar sonucunda meydana gelen kronik hiperglisemiyle karakterize metabolik bir hastalıktır. Karbonhidrat, yağ ve protein metabolizmasındaki düzensizliklerinden dolayı diyabetin etiyolojisi farklılıklar göstermektedir (Alberti ve ark., 1998; Abou-Seif, 2004). Diyabetin Tip I ve Tip II olmak üzere iyi bilinen iki tipi bulunmaktadır. Diyabet tiplerinden bir tanesi insüline bağımlı diyabet olarak adlandırılan ve genellikle genç yaşlarda görülen Tip I diyabettir. Tip I diyabet, pankreastaki beta hücrelerinin viral yollarla yada otoimmün sistemlerce hasar görmesi sonucu insülin üretme yeteneklerini yitirmeleri sonucu dışarıdan insülin tedavisi gerektiren diyabet tipidir. Tip II olarak adlandırılan diğer bir diyabet tipi ise insülinden bağımsız genellikle yetişkin insanlarda görülen diyabet tipidir. Tip II diyabette, insülin salgısı normal veya normalden fazla olmasına rağmen, glikozun hücre içerisine alınmamasıdır. Her iki diyabet tipinde de başlangıç sebepleri farklı olmasına rağmen hastalığın seyrinde, insülin sekresyonunda
ya da insülin hormonu duyarlılığında veya her ikisinde birden bozukluklar olabilmektedir (Cengiz ve Cengiz, 2000; Kazkayası, 2011).
2.4.1. Deneysel Diyabetik Hayvan Modelleri
Diyabet, günümüzde yaşam koşullarının etkisi ile gittikçe daha çok rastlanan ve bu sebepten dolayı tedavisi ile etki mekanizmaları konusunda çok çalışılmış ve hala çalışılmakta olan bir patolojidir. Bu konudaki çalışmalar öncelikle deney hayvanları üzerinde yapılarak elde edilen veriler insanlardaki uygulamalarda kullanılmaktadır (Szkudelski, 2001).
Deneysel diyabetik hayvan modelleri; hastalığın patogenezini, komplikasyonlarını ve diyabet riskini artıran genetik ya da çevresel etkileri anlamak ve çeşitli terapötik ajanların etkinliklerini değerlendirmek amacıyla kullanılan temel araçlardır.
Deneysel diyabetik hayvan modelleri spontan olabildiği gibi kimyasal, diyet, cerrahi operasyon ya da bunların kombinasyonları ile de elde edilebilir (Srinivasan ve Ramarao, 2007). Diyabeti taklit eden çok sayıda deneysel hayvan modelinin içinde en yaygın kullanılanları alloksan ya da streptozotosin (STZ) enjeksiyonu ile oluşturulan modellerdir. STZ, Streptomyces acromogenes’den izole edilen geniş spektrumlu bir antibiyotiktir. Antibiyotik olmasının yanısıra antilösemik ve karsinojen olduğu da gösterilmiştir. STZ’nin diyabetik etkisi, pankreas β hücrelerinin tahrip edilmesine dayanmaktadır (Szkudelski, 2001). STZ glukoz oksidasyonunu bozmak suretiyle insülinin biyosentezini ve salgılanmasını azaltmaktadır (Bedoya ve ark., 1996; Özer ve Gönül, 2006).
STZ, pankreas adacıkları içerisindeki beta hücreleri üzerindeki direkt sitotoksik etkisi ile hiperglisemiye neden olmaktadır. STZ’nin beta hücrelerinin ölümüne katkıda bulunan mekanizmaları arasında serbest radikal oluşumu gösterilmektedir. Ayrıca STZ’nin neden olduğu DNA dizisindeki kırılmalar ve alkilasyon da hücrelerin ölümüne neden olmaktadır (Srinivasan ve Ramarao, 2007).
2.4.2. Diyabet ve Oksidatif Stres
Son yıllarda yapılan çalışmalarda, diyabette gelişen komplikasyonların sebebinin artan oksidatif strese bağlı olduğu bildirilmektedir (Baynes, 1991; Bonnefont-Rousselot ve ark., 2000). Oksidatif stres, diyabet ve diyabetin daha sonraki komplikasyonlarında etkin olarak rol almaktadır. Bununla beraber diyabette oksidatif stresi artıran mekanizmalar olarak antioksidan savunma sistemindeki çeşitli değişikliklerin yanında, enzimatik olmayan glikozilasyon, otooksidatif glikozilasyon, sorbitol yolu aktivitesi, hipoksi gibi nedenler gösterilebilir (Brownlee, 2001).
Daha önceden yapılan çalışmalar, diyabette lipit peroksidasyonunun ve protein karbonillerinin normalden çok daha fazla oduğunu göstermiş ve bunun sebebini ise diyabette var olduğu düşünülen oksidan stresi ile açıklamışlardır (Baynes ve Thorpe, 1999; Coudray ve ark., 1999; Cho ve ark., 2002; Anwar ve Meki, 2003).
Yine günümüz literatür bilgileri diyabette var olduğu düşünülen oksidatif değişmelerin nedenlerinden birinin dokularındaki antioksidan enzim aktivitelerindeki değişiklikler olduğunu göstermektedir. Oksidatif streste, organizmanın antioksidan savunma sistemini oluşturan enzimlerin adaptif cevap ile uyarıldıklarını ve ayrıca, oksidatif stres karşısında enzim inaktivasyonunu gösteren çalışmalarda mevcuttur (Kakkar ve ark., 1995; Dinçer ve ark., 2002; Seven ve ark., 2004).
Diyabette protein glikozilasyonu ve glukoz otooksidasyonu sonucu serbest radikallerin oluşumu arttığı tespit edilmiş ve diyabetin kronik komplikasyonlarının etyopatogenezindeki artmış oksidatif stresin önemli rol oynadığı belirlenmiştir (Baynes ve Thorpe, 1999).
Diyabette, proteinlerin enzimatik olmayan yollarla glikoz bağlanması olayına glikozilizasyon denir. Bu olay diyabette ciddi bir artış gösterir. Oluşan bu glikozillenen proteinlerin oksidasyonu sonucu serbest radikaller meydana gelmektedir (Cengiz ve Cengiz, 2000; Akgül ve ark., 1999).
Diyabette artmış olan serbest radikaller genetik mutasyonlara, proteinlerde yapısal veya fonksiyonel bozukluklara sebep olmakla beraber lipitler, proteinler ve nükleik asitlerle etkileşerek membran bütünlüğünün kaybına da yol açmaktadır (Şekil 2.18) (Vincent ve ark., 2004; Memisogullari ve ark., 2003; Cherubini ve ark., 2005; Memişoğulları ve Bakan, 2004).
Şekil 2.18. Serbest radikal zararı sonucu ortaya çıkan ürünler
Yapılan bir çalışmada oksidatif stresin antioksidan enzimlerin mRNA ekspresyonlarında değişikliklere yol açtığı gösterilmiştir (Rohrdanz ve ark., 2001) Diyabetik hayvanların böbrekleri ile yapılan bir çalışmada, diyabette artan SOD ve GPx aktivitelerine paralel olarak bu enzimlerin mRNA seviyelerinin de arttığını, fakat buna tam ters olarak artan KAT aktivitesine karşı azalan KAT mRNA seviyesi bulunmuş ve bulunan bu aktivite düşüşünü KAT enziminin post-translasyonal seviyedeki değişikliğine bağlanılmıştır (Limaye ve ark., 2003).
Diyabette antioksidanların ve/veya oksidanların dokuların yapı ve fonksiyonu üzerine etkilerini düzenleyen mekanizmaların tam olarak aydınlatılamadığı bilinmekte ve birçok bilim insanı tarafından ileri çalışmaların yapılması önerilmektedir. Bu nedenle, oksidan stres kaynaklı patolojilerin moleküler seviyede kaynaklarının bulunması günümüzün önemli araştırma konuları arasındadır
2.5. Protein Fosforilasyonu
Aktivitesi bilinen birçok enzim, kovalent modifikasyonlarla özellikle de enzimlerin tireonin, tirozin ve serin gibi belirli birimlerine fosfatın eklenmesi veya çıkarılması ile düzenlenmektedir. Bu işlemler sonucu enzimin türüne ve aktivitesine göre fosforlanmış şekli, enzimin fosforlanmamış şekline göre daha fazla veya daha az aktif olabilmektedir. Yapılan çalışmalarda glikojen fosforilazın (glikojeni yıkan enzim) fosforlanması sonucu aktivitesi artarken, diğer taraftan glikojen sentaz’a (glikojen sentezleyen bir enzim) fosfat eklenmesi ile aktivitesinin azaldığı görülmüştür. Enzim aktivitelerini düzenlemek için gerçekleşen protein fosforilasyonu protein kinazlar, defosforilasyonu ise protein fosfatazlar tarafından yapılmaktadır. Hücre içinde önemli enzimatik kontroller protein tirozin kinaz (PTKs) ve protein tirozin fosfatazların (PTPs) farklı aktiviteleri ile gerçekleşmektedir.
Hücresel redoks durumu (indirgenme-yükseltgenme potansiyeli) serbest radikaller ve antioksidan sistemlerin birlikteliği ile belirlenmektedir. Hücresel redoks durumu hassas birçok protein ve/veya gen tarafından hissedilmekte ve bu redoks duyarlı sinyal proteinleri, hücresel metabolizmayı doğrudan kontrol ederek oksidan sinyalleri hücre içerisinde, mantıklı ve uygun hücresel yanıtlara çevirebilmektedir. Hücre içinde bulunan en önemli redoks duyarlı sinyal proteinleri protein kinazlar, protein fosfatazlar ve transkripsiyon faktörleridir (Surh ve ark., 2008).
Son yıllarda yapılan çalışmalar ile redoks sinyal iletimi ve protein fosforilasyon kararlı durum düzeyleri arasında güçlü bir ilişki olduğunu göstermiştir. Serbest radikal ve türevlerinin (özellikle hidrojen peroksitin) hücresel fosforilasyon düzeyini etkilediği ve sonuçta redoks duyarlı enzimlerin örneğin antioksidan enzimlerden KAT ve GPx’ın aktivite ve ekspresyonlarının değişebildiğini göstermiştir. Bununla birlikte birçok enzim ve proteinin gen ve protein ekspresyonunun antioksidanlar, oksidanlar ve hücresel oksidasyon-redüksiyon (redoks) potansiyeli (Sen ve Packer, 1996; Hardie, 1999; Watai ve ark., 2007; Koç ve Üstün, 2008; Surh ve ark., 2008) durumu tarafından düzenlendiği belirtilmiştir. Aynı zamanda bazı çalışmalar, redoks duyarlı proteinlerin doğrudan veya dolaylı olarak serbest radikaller tarafından okside olduğunu da göstermiştir (Şekil 2.19) (Storz ve Imlay, 1999; Halliwell ve Gutteridge, 2007).
Şekil 2.19. Oksidatif NF-κB aktivasyonu için oluşturulmuş model
Hidrojen peroksitin ve diğer oksidan stresi kaynağı moleküllerin, hücre içi PTP’lerin aktif bölgelerinde bulunan sistein gruplarını geri dönüşümlü olarak oksitlemesi sonucu hücrelerdeki toplam fosforolisyon düzeyinin değişebileceği ve ayrıca çeşitli hücresel antioksidanların da PTP gibi oksitlenmiş redoks duyarlı proteinlerin biyolojik aktivitelerini yenileyici fonksiyonları belirlenmeye başlanmıştır. Protein tirozin fosfatazların (PTPs) aktif bölgelerindeki tiyol gruplarının ve bunların katalitik bölgelerindeki sistein gruplarının tersinir oksidasyonu ile inaktivasyonu (Şekil 2.20) sonucu hücresel sinyal iletimi ve hücrelerde bulunan protein ve enzimlerin toplam fosforlanma düzeyinin etkilendiği daha önceki çalışmalarda da gösterilmiştir (Denu ve Tanner, 1998; Mahadev ve ark., 2001).