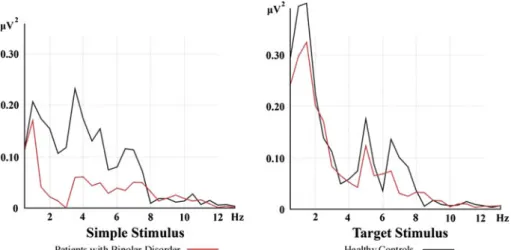
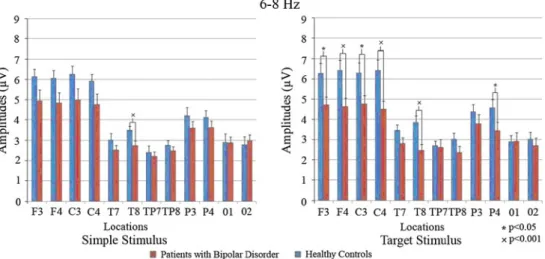
Decrease of Theta Response in Euthymic Bipolar Patients During an Oddball Paradigm
Tam metin
Şekil
Benzer Belgeler
Confirming these studies, we found that patients with bloodstream infections, gram-negative bloodstream infec- tion, and mortality due to FN had the AB/BB genotype more often
Clinical samples from four diseased chickens were examined for the detection and genotyping of IBV by virus isolation, a commercial real time reverse transcription polymerase
Çok de¤iflkenli analizlerde preterm eylem riskini etkileyen de¤iflkenlerin; genifl aile tipi, resmi nikâhl› olmama, preterm do¤um öyküsü, önceki gebeli¤inde erken
Gerekli sıkıĢtırma iĢleminin baĢarıyla uygulanması için, en uygun sıkıĢtırma aletinin tespit edilmesi son derece önemli olup yaygın olarak kullanılan baĢlıca
The main reason for the speed up is the initial partitioning of the output diffraction pattern (hologram) into segments, and then approximating the contribution of each object point
Önerme: “TTK’ya göre, finansal tabloların, Kamu Gözetimi, Muhasebe ve Denetim Standartları Kurumu’nca yayınlanan, Uluslararası Denetim Standartları’yla
EBCPG provides a comprehensive way to assist clinicians in making decision according to the visualized clinical practice guidelines while
[r]
