Carbonic anhydrase inhibitors: purification and inhibition studies of pigeon (Columba livia var. domestica) red blood cell carbonic anhydrase with sulfonamides
Tam metin
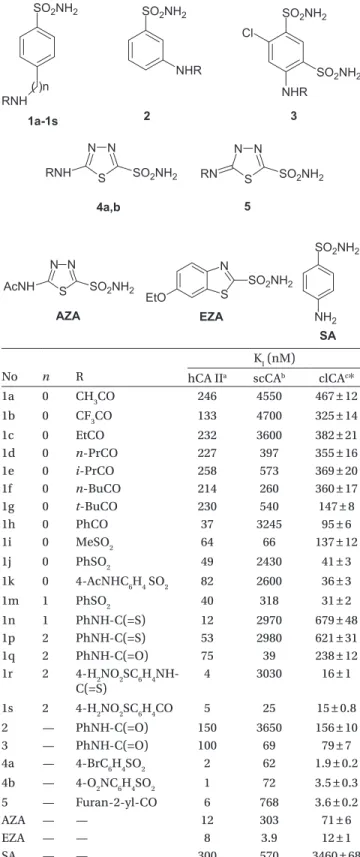
Şekil

Benzer Belgeler
Samsun ilinde alabalık üretimi (ağ kafes ve karadaki havuzlarda) yapılan işletmelerin yapısal ve ekonomik analizinin yapıldığı bu araştırmada; kapasite kullanım
Bu çalışma Bolu ilindeki çift toynaklı (Artiodactyla) memeli türlerinin belirlenmesi zamansal ve mekansal bilgilerinin ortaya konulması için yapılmıştır.. Bolu
Alanda tespit edilen Boz ayılar (a: Üretim faaliyeti öncesi, b: Üretim faaliyeti sonrası, c, d: beslenme amaçlı alanı kullanması).. Alanı sonbaharda daha yoğun
these coefficients will give a method which can be used in normality test of the data. In order to construct these intervals, it is required to define sampling distributions of
Methods For 70 normal eyes of individuals with varying age, the ETDRS chart acuity was determined as well as discriminated target perimetry using high-contrast targets presented for
32-Kullanmış olduğum telefonu kamera özelliği için tercih ettim 0,000 33-Kullanmış olduğum telefonun mp3 çalar özelliğinden dolayı tercih ettim 0,000 34-Kullanmış
tar sepsisi nedeni ile izlem ve tedavisi yap›lan hasta- larda literatürde bildirilen haz›rlay›c› etkenlerden; genifl etki alanl› antibiotik kullan›m›, bakteriyel sep-
Örnek alanlarda yol platformu, yol dışı (yol altı ve yol üstü) ve kontrol noktalarında tespit edilen yaban hayvanı görüntü sayısı arasında yapılan varyans
