Başlık: Köpek gençlik hastalığı virusu ile doğal olarak enfekte köpeklerin serebellumunda indüklenebilir nitrik oksit sentaz ekspresyonu Yazar(lar):YARIM, Murat; KARAYİĞİT, M.Önder; GACAR, Ayhan; KABAK, Yonca B.; GÜLBAHAR, M.Yavuz; GÜVENÇ, Tolga; SÖZMEN,
Tam metin
Şekil
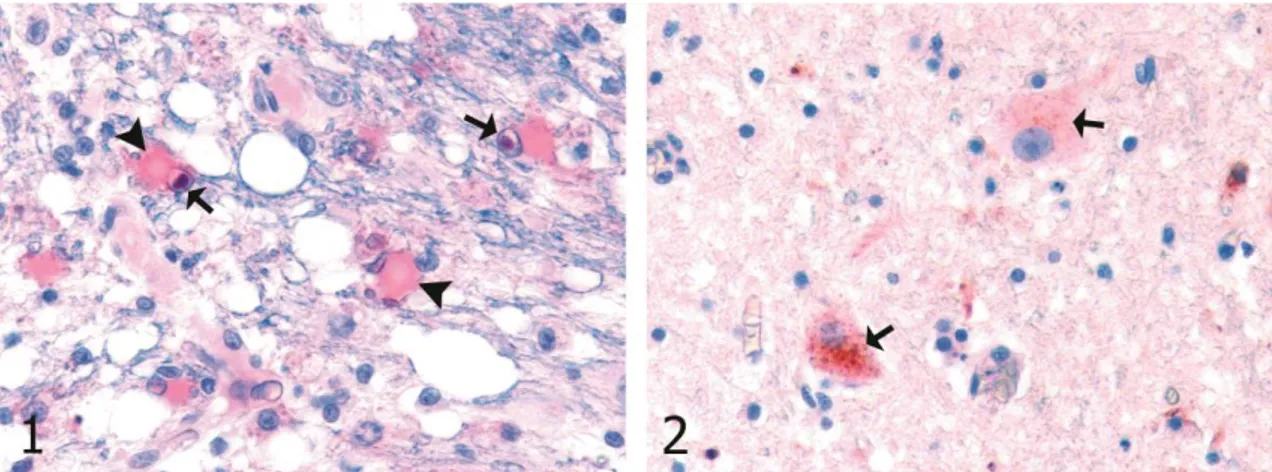

Benzer Belgeler
雙和醫院單孔達文西機器手臂,可一洞完成「子宮肌瘤、卵巢腫瘤、內膜癌」 等手術治療
雙和醫院器捐牆揭幕,謝謝您的大愛 雙和醫院為了推動器官捐贈的理念,並配合 6 週年院慶,於 6 月 14
雙和醫院圖書室榮獲「101 年度 NDDS 醫院圖書館類別服務件數」第 2 名 中華圖書資訊館際合作協會 2013 年 5 月 23 日假國立中山大學舉 辦第
We test whether inhibition of inducible nitric oxide synthase (iNOS) can exert a cytoprotective effect on cerebral endothelial cells upon stimulation by pro-inflammatory
Bu çalışmada, TNS tekniği kullanılarak uzantıları kesişen pist konfigürasyonundaki pistler için RNAV rota yapısına dayalı ve RNAV STAR olarak da tanımlanan geliş
Nitric oxide de- ficiency, which is also known as endothelial dysfunction, is the first step for the occurrence of many disease states in cardiovascular sys- tem including
S15. Çözümlenmiş olarak verilen sayıları yazalım.. S1."Yalan yanlış sözlerle beni kandırdı. " S1.Aşağıdaki doğa olaylarından hangisi sesin şiddeti-
Đ lköğretim sosyal bilgiler dersinde de sürece dayalı, öğrenciyi çok yönlü geliştirmeyi ve gelişimini izlemeyi hedefleyen değerlendirmeler için, bilinen
