Aktinomiset genomlarında yeni glikopeptid bileşiklerin genetik olarak taranması
Tam metin
Şekil


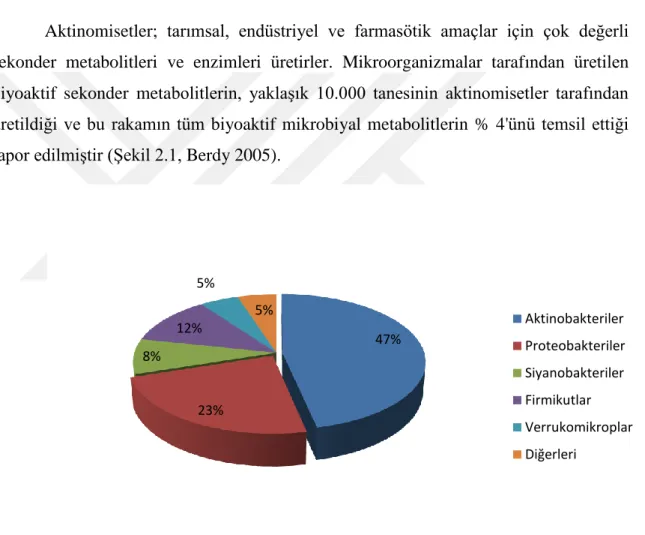



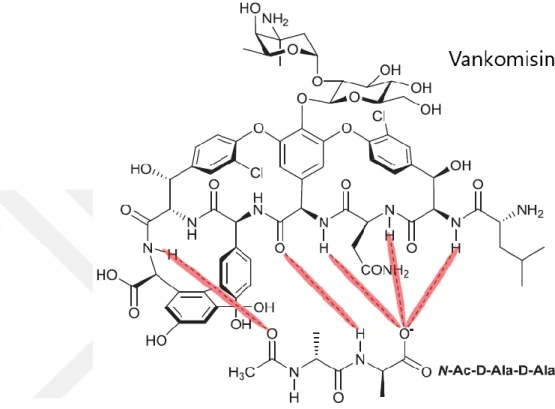


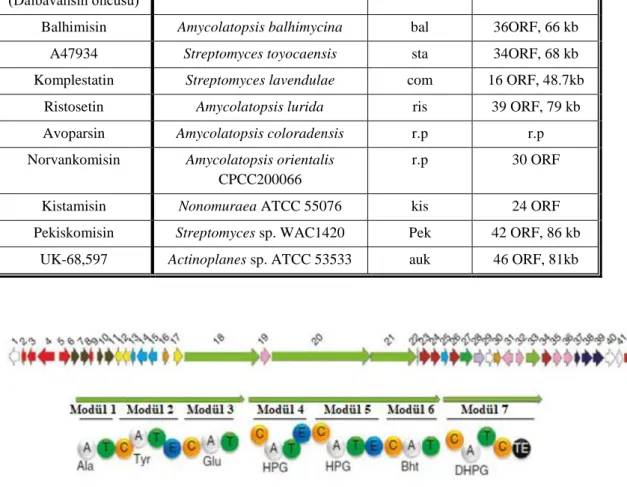
Benzer Belgeler
Ancak şunu rahatlıkla söyleyebilirim ki, böyle olağanüstü bir yapıtı yarata bilen bir toplumun insanı olmaktan o gece büyük gurur duydum.. Fazıl Say müzik
備急千金要方 針灸 -足三陰三陽穴流注法第二(下) 原文
Gerek bir embriyonun genetik yap›s›na dair bilginin nas›l kullan›laca¤›na, gerek- se klonlaman›n hangi koflullarda kabul edilebilir oldu¤una dair yasalar›n
Characterization of the 2-hydroxy-acid dehydrogenase McyI, encoded within the microcystin biosynthesis gene cluster of Microcystis aeruginosa PCC7806. Inactivation of an
For fast moving spare parts, Dilworth (1992) proposed many inventory control systems using normal distribution to approximate the demand during lead time.. In addition, Vereecke and
Sonuç olarak kişilik profili ve örgüt kültürü alt boyutları birlikte modellendiğinde kişilik profili alt boyutlarından; dışadönüklük, yumuşak
Topology optimization can be used to reduce the weight of existing knuckle component by 11% while meeting the strength requirement, with limited design space given with or
As the results of this study, it shows that Social Media Marketing activities have a significant effect on Brand Equity of healthy food products and Customer's Trust on the
