Farede siyatik sinir hasarı sonrası aquaporin-1 ekspresyon düzeyinin ölçülmesi
Tam metin
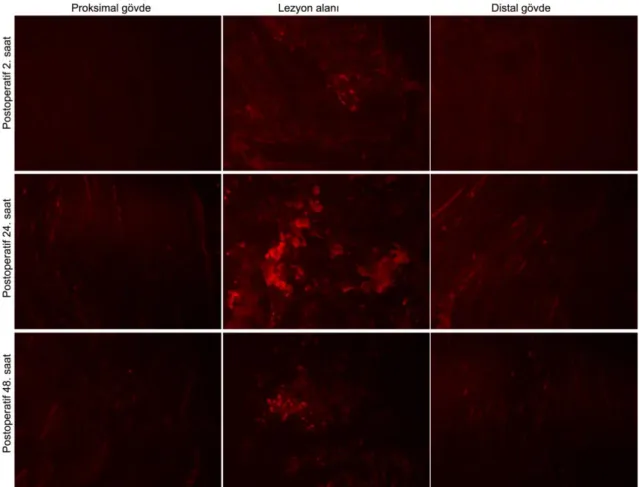
Şekil

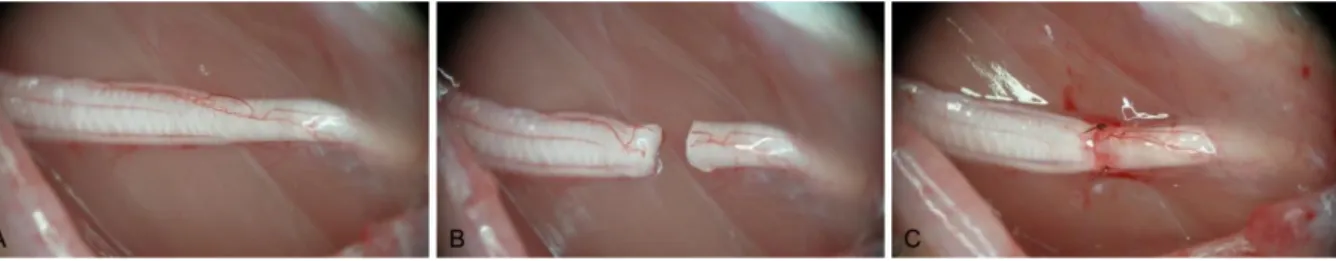
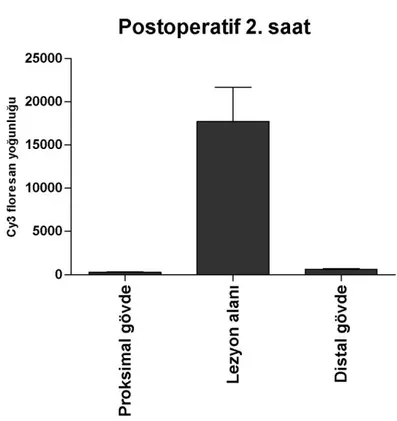
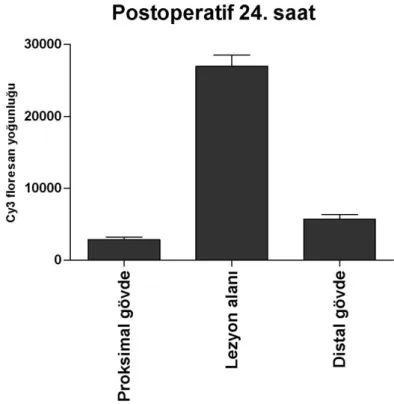

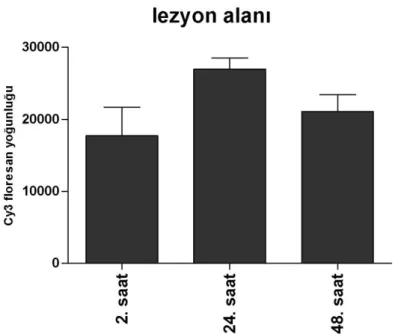
Benzer Belgeler
hücreleri tarafından merkezi sinir sisteminde oligodentroglia tarafından yapılır... Sinir Hücresinde Aksiyon
Tek bir işlevi vardır: göze abduksiyon yaptıran kası (m. rectus lateralis) innerve etmek..
Dejerine- Klumpke paralizisi (Alt brakial pleksus hasarı).. motor..
Motor Nöronlar (Efferent Nöronlar); SSS den kaynaklanıp kaslara, bezlere ve diğer nöronlara impuls götürür. Somatik motor nöronlar : İskelet kaslarını innerve
Sinir lifleri miyelinsizdir, sonlanmadan önce çevre bağ dokusu içinde sinir ağları yaparlar.. Duyuları
Sonuç olarak; hastanın sol karotid arter servikal segment distalinde görülen dissekan anevriztamik görünüme sekonder okülomotor sinir felci olduğu düşünülmekle beraber, travma
Bu çalışmada ROCK-1/LIMK-2/Kofilin sinyal yolağının bilate- ral CN hasarı yapılmış erkek ratlardaki korporal fibrozis ve ED üzerindeki etkisi incelenmiştir.. Ağırlığı
Bu çalışmamız ile primer tamir edilme şansını kaçır- mış olan ve tedavi sonrasında iyileşme imkânı olmamış yüksek seviyeli radyal sinir yaralanmalarında
