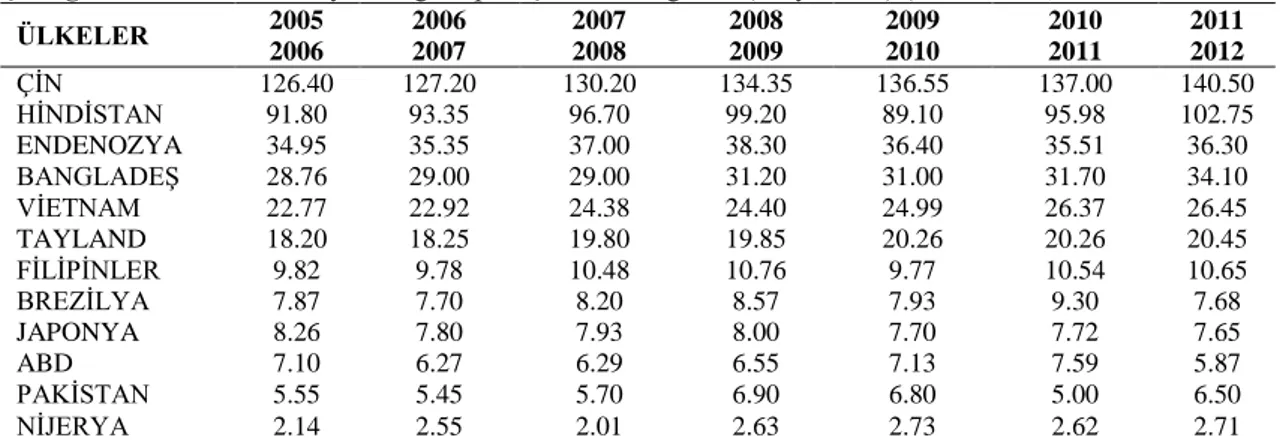
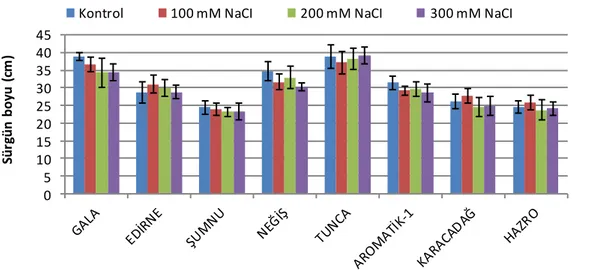
Tuz stresine maruz bırakılan çeltik (Oryza sativa L.) çeşitlerinde radikal söndürme ve antioksidan enzim aktiviteleri
Tam metin
Şekil








Outline
Benzer Belgeler
Ölçeğin Türkçe formunun ilköğretim öğrencileri ne uygulanması sonucunda elde edilen bu bulgular Coopersmith Özsaygı Envanteri'nin Türkçe sürü münde
Üniversite eğitimine yönelik talep ile ilgili olarak belirtilmesi gereken noktalardan biri de, söz konusu talebin, "olabildiğince yüksek düzeyde eğitim alarak daha
Nüfusun artış eğilimi göstermesi dışarıya olan göçlerin azalması ve Karahallı ilçe merkezine 1960 yılında elektrik gelmesi 1967 yılında ise elektriğin
İçe gömülü DMSG’nin momenti, generatör hızına göre uygun bir şekilde kontrol edilmektedir ve böylece rüzgâr hız algılayıcısı olmadan istenilen MGNT kontrolü
Betonda 28 günlük basınç dayanımı kaybı olmadan beton içindeki çimento miktarı, gerilim şiddeti, elektrotlar arası uzaklıklar değiştirilerek optimum en
Ekip, “kinon” (quinone) denen bir bileflim grubuna ait oldu¤u için “blatellaquinone” ad›n› verdi¤i feromonu yal›tmak için, gaz kromatografisi denen tekni¤in yeni
Ancak mayısa kadar şimdiki günlük miktar olan 63 bin metremikâbı tedricen art tırılacağından nisan ayında günlük miktarın 65 bin metre- mikâbını
Sonuçlar meme kanse- ri, kolon kanseri, kron hastalığı veya kalp krizi gibi çok ciddi hastalıklara yakalanma riski yanında göz rengi, laktoz toleransı veya dış kulak
