A quick and efficient method for DNA isolation from freshwater ostracods
Tam metin
Şekil
Benzer Belgeler
The findings of the present study on Turkish EFL learners‘ perception of teacher‘s authoritative roles may result from Turkey‘s education system in which the teacher has mostly
1234 Atiye ADAK ÖZDEMİR – Burcu ÖZDEMİR BECEREN Öğretmenlerin çocukların hangi çocuk kitabı yazarları daha çok sevdiği sorusuna ise çoğunlukla yazar içermeyen
Görüldüğü gibi Sepetçioğlu, Çatı adlı romanında, Osman Beyin kendi ça- ğındaki bireysel yaşam serüvenini yatay boyutta; tarihsel olan ile zamanla
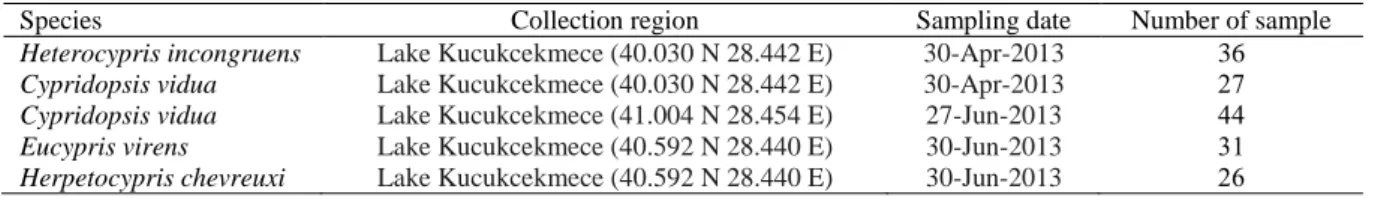
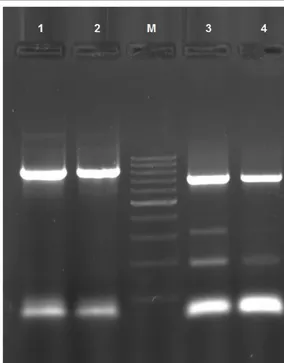
PCR (Polymerase chain reaction) technique is designed by Kerry Mullis in 1987. PCR is the in vitro amplification of a specific DNA part by primers. This technique is
After that, we proposed an enhanced version of proposed protocol, which satisfies mutual authentication with reader authentication against stronger adversaries, achieves destruc-
Attila Ilhan ‘m di zeleriyle tanışanlar aşk’ı daha anlamlı, rom an larını oku yanlar insanları da ha boyutlu, deneme leriyle gençliğinde tanışanlar
Kanüllü başsız vida grubunda ameliyat öncesi halluks valgus açısı, intermetatarsal açı ve distal metatarsal eklem açı ortalamaları erken ve geç ameliyat sonrası
Örnekleme alınan sağlık personelinin kadına yöne- lik şiddeti %95.7 oranında “kadına zarar veren fiziksel eylemlerdir” şeklinde tanımladığı (Tablo
