Glifosat bazlı herbisitin melanopsis praemorsa üzerindeki toksik etkileri
Tam metin
Şekil
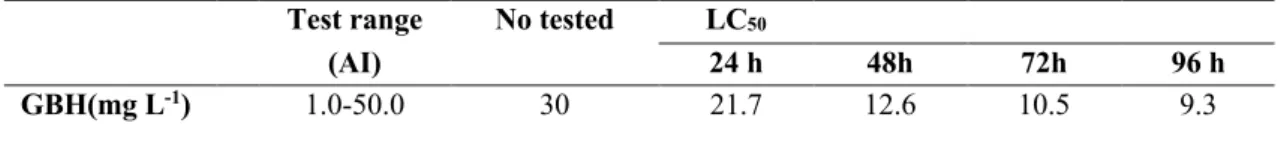
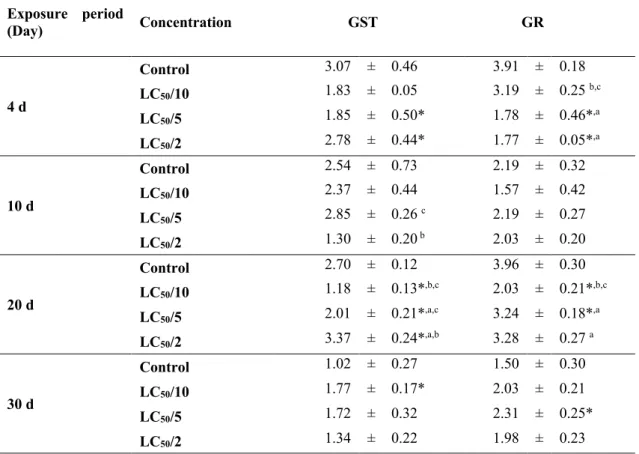
Benzer Belgeler
The adsorbent in the glass tube is called the stationary phase, while the solution containing mixture of the compounds poured into the column for separation is called
In this chapter, abolition of cizye (tax paid by non-Muslim subjects of the Empire) and establishment of bedel-i askeri (payment for Muslims non-Muslims who did not go to
Yet soon enough, he switches to “∞we∞”∞: “∞Il (a policeman) appelle divers noms et nous apprend que nous sommes soldats.∞” The meaning of “∞we∞”, however, changes
and because it was a nasty, nosy noise, they just drew noses for the N-sound, till they were tired (29); and they drew a picture of the big lake-pike’s mouth for the greedy Ga-sound
In this story, Joyce writes about the main character Eveline, whose desire to fit into the role of loyal Catholic daughter prevents her from fleeing with her sweetheart Frank to
Blunted pulmonary venous flow and re duced left atrial function has bee n deseribed in pati- ents with mitral stenosis but the effect of beta blackade on
• The Rashidun army was the primary military body of the Muslims during the Muslim conquests of the 7th century, serving alongside the Rashidun navy.. • The three most
Doğum ağırlığı, sütten kesme ağırlığı, anne sütü, ergin inek bedeni ile ilgili masrafların da dahil olduğu hayvan başına diğer hayvanlardan farklılığın dolar
