Salicylidene acylhydrazides attenuate survival of SH-SY5Y neuroblastoma cells through affecting mitotic regulator Speedy/RINGO and ERK/MAPK-PI3K/AKT signaling
Tam metin
Şekil
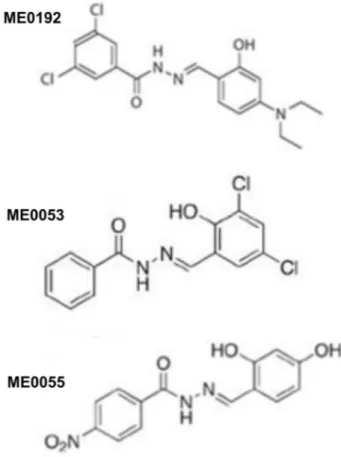

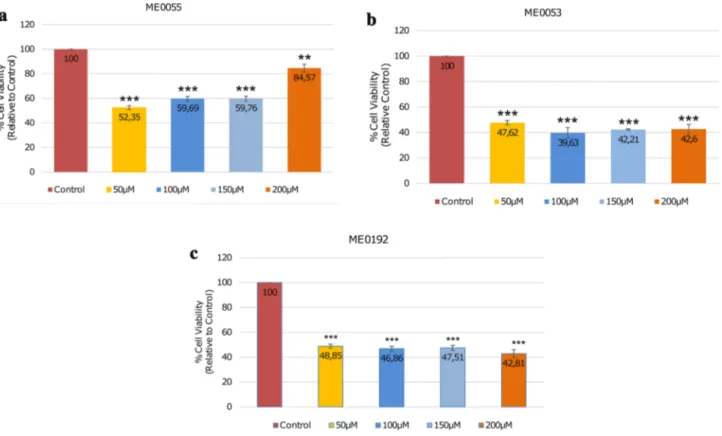
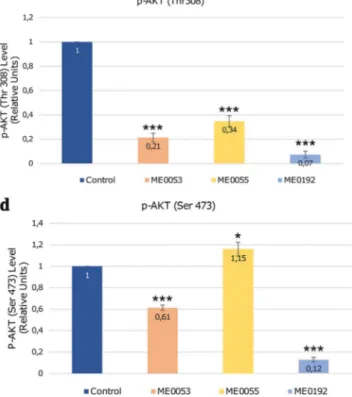
Benzer Belgeler
美好回憶、降低住院時的抗拒和焦慮感。【上圖:失智症專責病房溫馨的交誼廳 (左圖)及護理站的特殊紅磚瓦造型(右圖)】
雙和醫院婦產科溫國璋醫師貼心提醒,女性朋友忙碌之餘勿忽略婦科相關檢查
jik dışlanmaya maruz kalan üye sayısı arttıkça bireyin dört temel ihtiyacına (ait olma, benlik değeri, anlamlı varoluş, algılanan kontrol) yönelik tehdit
In this study, velocity profile, temperature profile, and corre- sponding Poiseuille and Nusselt number expressions are deter- mined for a fully developed gaseous flow in a
This study explored the effectiveness of commercial software in teaching grammar as compared to blended and teacher-led learning conditions, and the attitudes of students
For example, Özdamar and Canbolat’s (2018 , 23) research on Muslim Brotherhood leadership could only utilize twenty-six speeches and find 2,134 transitive verbs in to- tal from which
We found that coupling along the external E direction results to shift of the magnetic resonance frequency of the single SRR; the shift is upwards when the neighboring sides of the
Deneysel sonuçlar, Ba˘glamsal A˘gırlıklı Ortalamalı Tahminci (BAOT) ismini verdi˘gimiz bu algoritmanın sınıflar arası den- gesizlik olan veri kümelerinde AOT yönteminden
