Vet.Bil. Derg, (2004), 20, 4 :85-91
HASMER, HASAK VE ALMAN SiVAH
BAŞ
KOVUN
IRKLARıNIN
GENETiK
VAPILARININ MiKROSATELLiT MARKERLARLA
iNCELENMESi
Zafer Bulut
1@Mehmet
Nizamlıoğlu1In
ci
Togan
2A
nalys is o
f G
enetic
St
ructures
o
f H
asmer, Hasa
k and
Ge
rman
Bla
ck
Head Sheep
Bree
ds
by Using Mic
rosatellite
Markers
Özet: Buçalışmada, 1 saf (Alman SiyahBaş) ve 2 melez (Hasmer ve Hasak) olmak üzere 3farklıkoyunırkı arasındaki ge-netikfarklılıklbenzerlik araştırılmıştır.Bu amaçla,moleküler markeriardan 3 adet mikrosateütmarkeri (OarFCB20, OarJMP29 ve OarJMP58)kullanılmıştır.Irklararasıgenetikçeş~liliğin oldukça yüksekolduğuve FSTdeğerlerininistatistiki olarak önemli (P<O.OO1)olduğu belirlenmiştir. Hasmer ve Hasakırklan için hesaplanan FSTdeğerininistatistiki olarak önemsiz (P>O.OS)ol -duğuve buikiırkm genetikolarak birbirlerindenfarklı olmadıkları gözlemlenmiştir. Bütün populasyonlardaırkiçiFıs değerleri önemsiz (P>O.OS) olarakbulunmuşve Hardy-Weinberg dengesindensapmanın olmadığı gözlenmiştir.
AnahtarKelimeler : Koyun, GenetikYapı, Mikrosatell~
Summary: In this study, level of genetic diversity was investigated in one pure (German Black Head) and two crossbreed (Hasmer and Hasak) sheep breeds. For this purpose, three mcrosateüite markers (OarFCB20,OarJMP29 and OarJMPS8) were used.It was observed that geneticdiversity between these breedswere high and FST valueswere statistically significant (P<O.001).The FSTvalues ealculated for Hasmerand Hasak were found statistically insignificant (P>Ü.OS)and no genetic
dif-ference was observed betweenthese two crossbreeds. However,F
ıs
valueswere found statistieally insignificant (P>O.OS) and therewas no departure from the HardyWeinberg equilibrium in any of thesepopulations.KeyWords:Sheep, GeneticStrueture,Microsatell~e Gi riş
Türkiye'de,
çiftlik
hayvanlarıiçerisinde
%
59.3'lük bir dilimi
teşkileden (DIE 2
002)
ko
yun
sa-yısının%97 gibi büyük bir bölümünü yerli koyun
ırk ları,%3'lük bir bölümünü ise,
baştaMerinos ve
me-lezler
i
olmak
üzere
yabancıkoyun
ırkıarı oluşturmaktadır (Yalçın1990).
Türkiye'de Cumhuri
yetin
kuruluşundanbu
y
ana
koyun
ıslahıkonusunda özellikle melezlemeye
da-yanan birçok
çalışma yap ı lm ıştı r.Bunlar içerisinde
Merinoslaştırma,hem ilki
oluşturmaktahem de uzun
yıllarsürdürülen
ısrarlı çalışmalar,Türkiye'nin verimi
yüksek kültür
ırkı sayılabilecekbir
ırkıy
ani Türk
Me-rinosu
'nu
ortaya
çıkarmıştır(Tekin ve ark 2001).
Me-rinos
il
e yerli
ırklar arasındamelezlemeler
yapılaraki
ki
değişikTürk Merinosu tipi
geliştirilmiştir.Bunlar;
Karacabey Merinosu ve Orta Anadolu (Konya)
Me-r
inosu'dur
(Yalçın1990).
Çiftlik
hayvanlarındagenetik
çeşitlilik, doğalgen
kaynaklarının korunmasıgibi olgular giderek önem
ve ilgi
kazanmaktadır. Baştakoyun olmak üzere
diğerçiftlik
hayvanlarındada verim düzeylerini ar
-tırmak amacıyla yapılmaktaolan melezleme
ça-lışmalarıgenetik
çeşitliliğiazaltma
ternayülünded tr. Çoğunluklakültür
ırkıarının kalıtsaliçerikteki
payını ar-tırma doğrultusundaki .melezleme
çalışmaları,yerl
i
ırkıarıngenotiplerinin
giderek
azalmasıile
so-nuçlanmaktadır.Bu nedenle Türkiye'deki mevcut
ko
yun
ırkıarınıngenetik varyasyon niteliklerini
ayırıcıve özgül
yanlarınınklasik teknikler
dışındadaha
ay-rıntılımoleküler tekniklerle de belirlenmesine gerek
vardır(Soysal ve ark 2001).
Polimeraz Zincir Reaksiyonu (PZR)
tekniğinin geliştirilmesiylebiyoteknoloji
alanındason 20
yıli
çe-risinde hayal edilemeyecek
ilerlemeler
olmuştur. Farklı canlıtürlerine ait
populasyonlarınbireylerin
in
çeşitliDN
A
bölgeleri yükseltgenip inceleneb
ilmeye
başlanmıştır.Bir
t
ürün
populasyonlarıgibi evrimsel
olarak
yakın canlı gruplarının karşılaştırmalıolarak
ça-Iışılacağıya da populasyonlarda
yakınzamanda
oluş muşolan bir
olayınizlerinin
aranacağıbir durumda
GelişTarihi :18.10.2004 @:[email protected]
I.SelçukÜniversitesiVeterinerFakültesi.BiyokimyaAnabilimDalı .KONYA
2.OrtaDoğuTeknikUniversitesi, Biyoloji Bölümü,A KARA
*
Bu makaleSelçuk Universitesi BAP Koordinatörlüğü(200 1/097) tarafındandesteklenen "Türkiye' dekiBazı KoyunIrklannın GenetikYapıl annınMikrosatellitlerleIncelenmesi" isimlidoktora tezindenözetlenmiştir.BULUT. NlZAMLloGLU. TOGAN
en çok
kullanı langenetik
işaretlerdenbiri
de
mik-rosatellitlerdir (Togan ve ark
2003).
Mikrosatellitler,
ırkıarı tanımlamada kullanılanol-dukça uygun DNA
markerlarıdır(Loftus ve
ark 1999)
ve mikrosatellitlerin
yüksek polimorlik
yapıları(Gold-ste
in
ve Schlötterer 1998) koyun
ırkıarı arasındaönemli
farklılıklar olduğunu göstermiştir(Buchanan
ve ark 1994
,
Byrne
ve ark
(baskıda)).Türkiye
'de
DNA düzeyindeki
çalışmalaroldukça
yen
i
sayılabilecek durumdadır.Bu
çalışmadaDNA
'daki
3 mikrosatellit bölges
i
kullanılarak1 saf ve
2 me
lez
koyun
ırkındagenet
ik
yapı araştınlrruştır.Materyal
ve Meto
t
Çalışmada
1 saf (Alman Siyah
Baş)ve 2 melez
(Hasmer ve Hasak) olmak üzere 3
farklıkoyun
ır kındantoplam 60 örnek
kullanıldı.Kullanılan
hayvan materyal
i
içerisinde bulunan
Hasmer ve Hasak koyun genotipleri melezleme ça
-lı şmalarıyla
elde
edilmiştir.Konya Mer
inosu'nun
Alman S
iyah
Baş(ASB) ve Hampshire Down (HO)
elç
i
ırkıarıi
le me
lezienmesinden
Fl
'ler,
ASB Gl ve
HO Gl
'ler
elde
edilmiştir.Bu iki baba
hattındanelde
edi
len
melez/er
in
birbirinden
ayırtedilemeyecek
kadar
benzediğibi
ldirilmektedir.
Bu 2 baba
hattındangelen me
lezler
karıştırı larakdengeli bir tohumlama ile
Hasmer tip
i
elde
edilmiştir[HD(Fl) x HD(G1) x ASB
(F
l) x ASB (Gl
)
l.
Hasmer t
ipi
iç
in
açıklanangerekçe
ve
yaklaşımlari
le
ASB ve HD
'un
Akkaraman ile olan
m
elezierine
i
se Hasak ism
i
verilmiştir(Tek
in
ve ark
2
001).
Çalışma
materya
li
olarak kan
kullanılmıştır.B
i-rey
ler
seçilirken özell
ikle
b
ireyler
arasında yakın ak-rabalı k ilişkisi olmamas ınaözen
gösterilmiştir. Alınank
an örn
eklerinden
DNA'yl
izo
le
etmek için Standart
F
enollKloroform Yöntem
i
uyguıanmıştır.Araştırmada
genom
ik
DNA'da 3 mikrosate
llit
(
OarFCB20,
OarJMP29 ve OarJMP58) bö
lgesi
po
-Ii
meraz zinc
ir
reaks
iyonu
(PZR
)
yöntem
iyle
amp
lifiye
ed
ildi.
PZR
karışım ıher b
ir
örnek
i
çin son hac
im
15
LLLo
lacak
şekilde0
,5
ml'
lik
ster
il
ependorl tüplerinde
hazı rlanmıştır.MgC
I2
konsantrasyon
u
OarFCB20
10-k
usu
i
çin 4
.5
mM, OarJMP29 ve OarJMP58
l
okusu
içi
n 1
.5
'
mM olarak
ku
llanılmıştır
.
PZR
işlem
i
rad-yoakt
if
işaretlidNTP
(
a-
33P) kullanılarakger
-çekleştirilmiştir.
Pol
iakrilamit
je
l
elektroforez
i
(PAGE) ile PZR
ü
rünleri
değerlendirilerekalleller
in
fenotip
leri
be-li rlenmiştir.
PAGE
işlemi ,DNA d
izi
analiz
i
(Thermo
E
C 160) alet
i
ve 35 X 45 cm cam plaka
lar
ku
l-lanılarak
d
ikey
(vert
ikal)
olarak
gerçekleştirilmiştir.Jelin
y
ürütme
işlemitamamlandıktansonra
kro-motograf
i
kağıd ı(Whatman
3 MM
) üzerine
alınmıştır. Kağ ıtüzerine geçen jelin
yüzeyi
strech film
yar-dı mıyla kaplanm ışve vakumlu bir jel
kurutucu
(Ther-mo Savant
SGD2000
Siab
Gel Dryer)
kullanılarak80
°
C'de
20-25 dk
kurutulmuştur.Kurutma
işleminden sonra jel,
karanlıkodada
ışıkalmayan bir
metal kaset (Exposure Casette,
Sigma E-9510)
içe-risine
yerleşti rilmişve üzerine özel 35 X 45 cm eba
-dında
oloradyografi
filmi (Kodak Biomax MR-2
,
Sigma Z35,041-9)
yerleştirilmiştir. Kullanılanrad-yoaktif madden
in
yeni veya esk
i
oluşunagöre
j
eller
kaset içerisinde 2-5 gün süreyle
bekletilmiştir.Daha
sonra kasetler Selçuk Ün
iversitesi
Veteriner
F
a-kültesi
Cerrah
i
Anabilim
Dalı'ndabulunan otomat
ik
film banyosu
eihazında(AFP Imaging Min
i-Med/90)
karanlık
odada
açılarakbanyo
edilmiştir.Banyo
işlemindensonra je
l
üzer
inde
görüne
n
DNA
bantlarıDNA ladderlar
yardımıylanegatoskop
üzerinde okunarak her b
ir
mikrosatellit lokusuna ai
t
ailelerin fenotipleri (baz çifti
uzunlukları) aynıjel
üze-rinde yürütülen DNA
standardı(DNA Sequencing
Kit)
yardımıyla belirlenmiştirve böylece
çalışılanb
i-reylerin genotipleri
saptanmıştır.Bireylerin
çalışılanmikrosatellit
lokuslarınada
-yalı
genotipleri belirlendikten sonra
,
ve
riler
b
il-gisayarda
değerlendirilmiştir.Irk iç
i
genet
ik
çeşitlilik(varyasyon) düzeyleri
;
1
-
Göz
lenen
(Ho
)
ve
bek-lenen (He) heterozigotluklar
il
e, 2
-
Lokus ve
ırkiçin
gözlenen ve ortalama alle
l
sayılarıo
larak
v
e
Wright'ın
F istatistik
değerleriGENETIX 4.0
prog-ramı
(Belkhir ve ark
1
996-2000)
kullanılarakhe-saplanrruştır.
FS
T
değerleri ırklar arası farklılaşmayıölçmek iç
in,
Fıs değerleride
ırkiç
i
Ha
rdy-Weinberg
denges
inden
sapmayıö
lçmek
iç
in
kullan ı lmıştır.Fa
k-töriye
l
birleştiricianaliz
i
de
(Facto
rial
Co
r-respondence Ana
lysis,
FCA)
aynı yazılımku
l-lanılarak yapılmıştır.
GeneClass
programı kullanılarak YerleştirmeTest
i
(Assignmen
t
Tes
t)
yapılmış(
Cornuet and
Lu-ikart 1996
)
ve Popu
lations
ı.oi
statistik
programı(http://www
.cnrs-gif/pgelbioinfo/populations)
kul
-lanılarak ırklar arasında
Ne
i (
1987)'nin DA ge
netik
uzaklıkları belirlenmiştir.
Sa
itou
ve Ne
i (
1987)'nin
komşu birleştirme ağacı
(Neighbor
-Joining
T
ree,
NJT) yöntem
i i
le ç
izilen
dendrogramla bu
uzaklıklargörselleştirilmiştir.
Bulgular
Çalışmada,
toplam 3 popu
lasyon,
60 b
irey
ve 3
mikrosatellit lokusu
kullanılmıştır. Çalışılan3 m
ik-rosatellit lokusunda toplam 43 allel
gözlemlenmiştir.Bu allellerin 13 tanes
i
OarFCB20
,
18 tanes
i
Hasmer, Hasak ve Alman SiyahBaşKoyun •.•
OarJMP29 ve 12 tanesi OarJMP58
lokusundadırve
gözlemlenen aileller içerisinde 17 tanesi sadece tek
bir
ırkda görüldüğünden,o
ırkaözgü
(private
allel)'dür. Su özgün aileller 3 adet ASS, 6 adet Hasak
ve 8 adette Hasmer
ırkında gözlemlenmiştir(Tablo
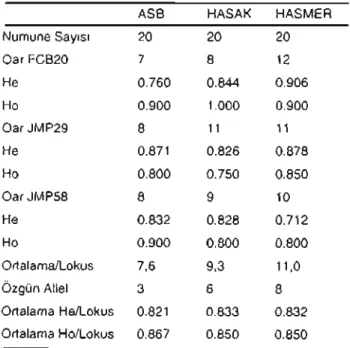
1).Tablo 1.trklardaGörülen GenetikÇeşitlilik
ASB HASAK HASMER
NumuneSayısı 20 20 20 Oar FCB20 7 8 12 He 0.760 0.844 0.906 Ho 0.900 1.000 0.900 OarJMP29 8 11 11 He 0.871 0.826 0.878 Ho 0.800 0.750 0.850 OarJMP58 8 9 10 He 0.832 0.828 0.712 Ho 0.900 0.800 0.800 Ortalama/lokus 7,6 9,3 11,0 ÖzgünAllel 3 6 8 Ortalama He/lokus 0.821 0.833 0.832 Ortalama Ho/lokus 0.867 0.850 0.850 He:Beklenen heterozigotluk, Ho:Gözlenen heterozigotluk değerlerinigöstermektedir.
,....- '...,u
Irklar için genetik
çeşitliliğin(Tablo 1) oldukça
yüksek
olduğu(Ho
=
0.850 - 0.867, He
=
0.821
-0.833)
gözlemlenmiştir.Irklar için hesaplanan genel
Fıs değerlerinin
-0.022 ile -0.057
arasında değiştiği belirlenmişve
Fıs değerlerininistatistiki olarak
"önemli"
olmadığı(P>0.05) (Tablo 2), böylece hiçbir
ırk
için Hardy-Weinberg dengesinden sapma
ol-madığı gözlenmiştir.
Tablo 2.FıS DeğerleriTablosu
Irk
FıSASB -0.057ns
HASAK -0.022 ns
HASMER -0.022 ns
ns :Istaiıl<Selolarakanlamlı aegıl
Irklar
arası farkınönemli olup
olmadığınıgör-mek için hesaplanan ikili
sonuçlarınagöre ASS
ır kının diğerlerinden farklıve bu
farkınistatistiki olarak
önemli (P<0.001)
olduğufakat melezleme
ça-lışmalarıyla
elde edilen Hasak ve Hasmer
ırkıarıkar-şılaştırmaları
sonucu elde edilen FST
değerininis-tatistiki olarak önemsiz (P>0.05)
olduğu(Tablo 3)
ve bu iki
ırkınbirbirinden genetik olarak
farklıol-madığı gözlemlenmiştir. Ayrıca
bütün
ırklar arasındagörülen ortalama FST
değeri0.072 olarak
bu-lunmuştur.Tablo 3. FST (Alt diagonal) ve DA (Üst diagonal)Değerleri Tablosu
ASB HASAK HASMER
0.459 ASB HASAK 0.113··· HASMER 0.087··· 0.010ns 0.322 0.185 r - - - -U \..'i: u . ' - - tl-\."".\IlM
Şekil 1.Komşu Birleştirme(NJT) Metodu Ile ÇizilenAğaç
(·P<0.05, ··P<0.01, ···P<0.001, ns : istatikselolarak an-lamlı değil)
Tablo 3'te bulunan üst diagonal
ırkıarınbir-birinden
uzaklıklarını DA değerlericinsinden
ver-mektedir.
DA değerlerinegöre en yüksek genetik
uzaklığın (DA =
0.459) Hasak ile ASS
arasında,en
düşük
genetik
uzaklığınise
(DA=
0.185) Hasak ile
Hasmer
ırkıarı arasında olduğuhem tablodan
(Tablo 3), hem de
DAgenetik
uzaklığı kullanılarakBULur, NtZAMLloGLU. TOOAN
,
A i('IM '! tl•
i ••
•
•
i
j•
,
•
•
,
_:.:--=~::~;-..-.::=-=-Şekil2.Factorial CorrespondenceAnalysis (FCA)Grafiği
Sarı:ASB, Beyaz:Hasmer,Mavi: Hasak
Tablo 4.Bazıevcil hayvanırkıarındamikrosatellitlerileyapılan çalışmalann karşılaştırılması
Tür Ismi Lokus He Ortalama AllelSayısı .Kaynak
Koyun 10 Adet 0.184- 0.732 2.4 - 4.4 Gutierraz-Espelata ve ark (2000)
Koyun 15 Adet 0.574- 0.760 4.4 - 10.9 Grigaliunaiteve ark (2003)
Koyun 3 Adet 0.726- 0.782 6.67 - 9 Soysal ve ark (2001)
Koyun 20 Adet 0.679- 0.763 Diez-Tasconve ark (2000)
Sığır 17 Adet 0.53 - 0.66 6 - 7.7 Moazami-Goudarzive ark (1997)
Sığır 20 Adet 0.54 - 0.79 Loftus ve ark(1999)
Keçi 20 Adet 0.51 - 0.58 Saitbekovave ark (1999)
Keçi 17 Adet 0.611 - 0.784 5.24 - 7.77 Li ve ark(2002)
Koyun 3 Adet 0.816- 0.864 7.6- 11 SunulanÇalışma
Ç
izilen
ağaca bakıldığında2 grup
oluşmuştur,bun-lardan Alman S
iyah
Baş(ASB)
ayrıbir grup
,
Hasak
ve Hasmer
i
se b
irlikte
b
ir
grup
oıuşturmuştur.I
rklar
arasındaki farkı yansıtanbir öncek
i
ağaçtan sonra
ırkıaraait b
ireylerin
ne kadar
ayrışabildiğinigörmek iç
in
Factoria
l
Correspondence Ana
lysis
(
FCA)
grafiği çizilmiştir (Şekil2)
.
FCA
grafiğinde(Şekil
2) ASB
ırkıbirey
lerinin
ayrılmış olduğu,Hasak
ve Hasmer
ırkıarınaait b
ireylerin i
se b
ir
arada
k
ü-melendiği gözlenmiştir.
N
itekim
Gene Class (1
.0.02)
programıkul
-lanılarak yapılan yerleştirme
(ass
ignment)
test
inde,
bireyler kendi
ırkıarından başka,en az 2
farklı ırkadaha dahil edilmektedir
.
Yani bireyle
r
çalışılan3
lokus
bakımındangenot
ip
olarak
kend
i
f
e-notiplerindek
i
ırkta gruplanmamıştı r.Tartışma
ve Sonuç
a- Irk Içi Genet
ik
Çeşitlilikve Hardy-We
inberg
Dengesinde O
lmak
Sunulan
çalışmada,3
m
ikrosatellit l
okusu
(OarFCB20
,
OarJMP29 ve OarJMP58) yönünde
n
ırkiçi ve
ırklar arasıgenet
ik
varyasyonu
n
(çe
-şitliliğin)
evcil hayvanlarda mikrosatellitlerle
yapı lança-Hasmer ,BIlSak ,,~AlmanStyahHa, Koyun...
hşmatardan
yüksek oldu{lu
gözlenmiştir.Yapılan bazı çalışmalarda
tespit edilen topl
am
au
et
sayılanna baklldlOında,Farid ve ark
(
2000)
1
0
koyu
n
ırkında10 m
ikrosatelllt lokusu
kullandıklança
-tışmada
t
oplam
93 ade
t
allel, Soysal ve ark
(
200 1) 3
m
ikrosatellit Iokus
uyla
Türkıye'deki5 k
oyun
ırkındayaptıklan çalışmada
ise
t
oplam
45 adet
aııeltespit
ettiklerin
i
bi
ldirmektedirler
.
Ayrıcabirçok
çalışmadaırkıara
O
zgün a
lleller (
p rivale
aıle/) belirtilmiştirve
k~yun
larda
yapılanbu
çanşmeıara baklldlQında;G
u-tie
rraz-Espelata
ve
ark
(
2000) 7 adet
O
zgün alle
l,
G
rigaliuna ite
ve ark
(
2003)
36
adet
O
zgün alle
l,
Ar
-ranı
ve ark (2
00 1a)
ise
15
a
det
O
zgün a
llel
tespit
et-t
iklerini bildi
rmektedirler. S
unulan
çalışmadaise 5
ırkda,3
mikro
satellil
böl
gesi nde toptam 43 adet
aıle!tespit
edilmişv
e
bu
a
ile llerden
17 tanesinin
O
z
gün
enet
ok1
u{1u
beürenmıştır. Çalışılankoyun
ırkıannm,di
Qer
araştıncılannbildi
rdiOi deOenere göre
daha
yO
ksek a
Jlel
sayısınav
e
ge
netik
çeşitlili{ıesa
hip
0'-duğu
g
Orü
l
mekt ed ir.
B
unun
en
O
nemli sebebin
in
Türkıye'nin
COOra
fi
bölge ol
arak koyunun eveill
ilme
kaynaQınayakınolmasıyla ilişkili olduğu şeklinde
dü-şünülmektedir (Yalçın
1986
,
Brut
ord
ve ark 2003)
.
N
itekim
çalışmada kullanı lanme
lez l
rk1ann ol
u-şumunadaha
ö
ncede belirtildi{l
i
gib
i
yerl
i
koyun
ırkIanmızında
g
enet ik
ola
rak
katkısı vardırve bu melez
ırklary
erli
ırklanmızaa
it
aüeüert de
taşımaktadırlar.Ozg
ün
a
llel
sayısının
ise yukardaki be
lirtilen
ça
-lışmalara
benz
er
d
eQerler
gOsterdiğiçöz
-lenmektedır.Fakat
ö
zqün
a
ilellerin
frekanslannın düşük olması(
0.0250
-
0
.1750)
alle
llerin
ırklarıbe-l
irleyen
ayırtaçlarola
rak
kullanılmasınai
mkan
ve
r-memektedi
r.
Gutterraz
-Espetata
ve ark (2000) koyu
nlarda
yaptıklan
b
ir
çalışmada,bü
tün
ırklarınH
ardy-We
inberg
(
H
W)
dengesind
e
olduklarınıç
öz-le
mlerken. Arranz ve a
rk
(1998)'nınk
oyu
nlarda
y
ap-tıklan
d
iOer
b
i
r
çalışmadaise
,
çalışılan ırklann bazıl
okuslar
açısındanHW
dengesındensapmalar çös
-terdiO
i
bildiıilmektedir. Fıs değeriak
rebah
yetiştirmekatsayısı
o
larak da
b
ilinmektedir
ve su
nulan
ça-lışmada ırk
içi HW denge
sinden
sapma
olmaması,s
eçilen b
ireylerin
yakına
kraba
olmamasındankay-naklanmaktadı r.
b-
l
rklar
v
e lrklarm B
ireyleri
ArasındakiGenetik
Farklılı klar
Irkıar arası
genetik
larklıl ıklannbe
lirlenmes i
için
hes
ap
lana
n F
ST
değerlerinegO
re aralannda fark
g
ö
zlenmeyen
lek
i
ki
li ol
an Ha
sak
ve Hasmer
me-l
ezlerinin her
ikis
i
nde d
e
Hampsh
ire
Down
ve Alman
S
iyah
Baş ırklannın katkılan bulLrlmaktadır.Bu
ne-de
nle
bu
i
kili birbi
rlerine ço
k benziyor olabi
lir
.
Yine
de
kullanılanm
ikrosate llit Iokusu
sayısıarttmlacak
ol
ursa
bu ikil
i
arasındad
a
f
ark gOrülme ihtimali yük
-sek
olacaktır.lrk1ann i
kili
karşılaştırmalansonucu
hesaplanan
F
ST deOerlerine gOre en küçük
deşer.H
asak ne Hasmer
arasındagOzle
nen
0
.0
10
Oıup,en
yüksek
deı':Jerise
AS
e
i
le Hasak
arasındagOzlenen
0
.113
deQerid
ir.
Koyun
larda
yapılandger
çalışmalardabulunan
o
rtalama FST de{)erlerin
i
GUlienaz
-Espelata
ve ark
(
2000) 0
.264,
Farid ve ark
(
2000) 0
.163,
B
yme ve
ark
(
basktda) 0
.
18
olarak tespit
etmişlerdir.Sunulan
çalışmada
bu
lunan o
rtalama F
ST
deOe
ri (
0
.
072)
bahsed
ilen
çalışmalardanoldukça
dUşüktür.O
halde
,
çalışmada kullanılan ırklarbirbirinden
farklıolsa da genet
ik
olarak
aralarındaki farklılaşmaA
v-rupa
ırklan arasındarastlanan düzeyde
deı':Jildir.Yu
-karda bahsed
ilen
GUlienaz
-Espelata
ve ark
(
2000)
il
e Farid ve ark
(2OOO)'nınçalışmalanndatö'ar
m
ik-rosatellit Iokusu
kullanılmıştır.Byme ve ark (ba
s-kıda)ise
20 m
ikrosatelJil Io
kusu
kullanmışlardır.Su·
n
ulan
çalışmadais
e ancak 3 m
ikrosate llit Io
kusu
çahşılabilmiştir. Araştırmada kullanılan
lrk1ann
rre-lezleri
i
le ge
nol ipik
olarak
yakınolmasıbeklenen b
ir
durumdur
.
Ayrıca çalışılan ırklar sınırlıbir COOrafyayl
tems
il
etmekted
irler
.
Büt
Ü
n
bu nedenlerden or
-talama
FST
deı':Jerinin dj~er araştmertanngöz
-lediklerinden daha
düşükolmasıbeklenme
lidir.
A
rranz ve ark
(2001b)'nın ıspanyolkoyun
ırkIan
nda
yaptıklan çalışmada,F
ST
deOeri 0
,068
olarak
belirlenmiş
ve
ıspanyolkoyun
ırklan arasında yakınbi
r
gene
tik
ilişkio
lduQunu
o
rtaya
koymuşlardır.Bu
so
nuç
yukarda ba
hsedildiOi
şekildeyerel bi
r
coğrafyayı
t
emsil e
tme ktedir
ve s
unulan
çalışmadakiF
ST
sonuçlarınıdes
tekle r n
itelikted
i
r
.
I
rk
la
r
arası farklılıklarF
ST deOerlerine gOre
i
s-t
atistiki olarak Onemli
olmalarınaraOmen
,
Irkla
r
b
ir-bi
rlerinden
kes
i
n
çizg
ilerle
aynfan
,
bireyleri b
irbirine
çok benzeye
n
O
zgün
daOıhmlarg
Os
termemekted
i
r.
B
u durum 3 boyutlu düzlemde FC
A
metoduna gö
re
çiz
ilen
gra
fikte
de
(Şekil2)
gOzlenmiştLA
rranz ve ark (2001b) koyunlarda 18 m
ik-rosatellit
Io
kusu ku
llanarak
yaptıklan çalışmada.b
i
-reylerin hang
i
ırkaa
it
olduklannıbe
linernek için y
er-leştirme(
assignme n
t
)
testi
n
i
ve
seçene
klerden
B
ayesian metodunu ku
llanarak
en
yü
ksek
pe
r-formansı
elde en
ikle rini
belirtmişlerdir.Sunu
lan
ça-Irşmada
da
aynıyöntem
kullan ı lmıştır.M
audet ve
ark
{20(2)'nın Fransız sı~ırlrk1a
nnda
yaptıkları çal ışmada,yerleştirmete
stinin
güven
ara-IıOl nın(
OrneOin
P
<O
,
05'1en O
,OOl'e)
ve lrk1ar ara
-sında benzerli~inin artmasıytat
est in
yerleştirmeg
ü-BULUT. NlZAMLIoGLU.TOGAN
cünün
azaldığını gözlemişlerdir.Sunulan
çalışmadada. Bayesian metodu
kul-lanılarak
3 mikrosatellit lokusuyla
yerleştirmetesti ya·
pılmış
fakat daha önce 3 boyutlu FCA
grafiğindegöz-lenen
örtüşen dağılımla oluşanbeklentimize paralel
olarak bireylerin kendi
ırkıarındatam olarak
grup-lanmadığı,
bir bireyin kendi
ırkından başkaen az 2
farklı ırkta
daha
gruplandığı gözlemlenmiştir.Bu
du-rumun Byrne ve ark
(baskıda)'dada
gözlendiğigibi
genel bir durum olmakla beraber, Maudet ve ark
(2002)'nın belirttiği
gibi
ırklar arasındakigenetik
fark-lılıkların
çok büyük
olmamasındanda
kaynaklandığıdüşünülmektedir.
Genellikle mikrosatellitlerle yurt
dışında yapılançalışmaların çoğunda
PCR
işlemindensonraki
elekt-roforez
işlemiDNA Genetik Analizatör
cihazlarındaotomatik olarak
tamamlanmışve bu sayede en az
15
-20
mikrosatellit
lokusuyla
çalışmalarya-pılabilmiştir.
Sunulan
çalışmadaPZR
işlemindenson-rak
i
elektroforez
işlemininmanuelolarak
ya-pılmasından dolayı
daha az mikrosatellit lokusu ile
çalışılabilmiştir.
Ancak,
çalışmaTürkiye'de DNA
mar-kerları
ile
yapılan çalışmalar arasındailkierden birisi
olmasıyla
önem
kazanmaktadır.Bu alanda bundan
sonra planlanan
çalışmalarayol göstermesi ve veri
tabanı sağlaması açısından
önemlidir.
Sonuç olarak,
ırkıardagörülen genetik
çeşitliliğinoldukça yüksek
çıktığı(Ho= 0.850 - 0.867 ve He=
0.821 - 0.833)
,
çalışılan ırkıarınömeklerinin
Hardy-Weinberg dengesinde
oldukları, ırkıarınikili
kar-şılaştırmaları
sonucunda bulunan FST
değerlerininis-tat
istiki
olarak önemli (P< 0.001) ancak
farklılaşmadüzeylerinin
diğerbirçok koyun
ırkı arasındagö-rülenden az
olduğu belirlenmiştir.Sadece melezleme
çalışmalarıyla
elde edilen Hasak ve Hasmer
ırkıarının karşılaştırmalarısonucu elde edilen FST
değerininis-tatistiki olarak önemsiz
olduğuve bu iki
ırkıngenetik
olarak birbirlerinden
farklı olmadıkları gözlenmiştir.Kaynaklar
Arranz JJ, Bayon Y, Primilivo FS (1998) Genetic
re-lationships among spanish sheep using microsatellites.
Anim Genel,29(6):435-440.
Arranz JJ, Bayon Y,Primitivo FS (2001a) Genetic variation
at microsatellite loci in Spanish sheep. Smail Ruminant Re-search; 39(1):3-10
Arranz JJ. Bayon Y, Primitivo FS (2001 b) Differentiation
among Spanish sheep breeds using microsatellites Genet
Sel Evol ;33(5):529-42.
Belkhir K,Borsa P.Chikhi L, Goudet J and Bonhomme F (1996-2000) Genetix 4.0 WindowsTM software population
genetics, Laboratoire Genome, Populations, lnteractions, University of Montpellier. France.
Bruford MW, Bradley DG, Luikart G (2003) DNA markers reveal the complexity of livestock domestication, Nat Rev
Genet., 4(11),900·910.
Buchanan FC, Adams
LJ,
Liltlejohn RP, Maddox JF andCrawford AM (1994) Delennination of Evolutionary Re-lationships Among Sheep Breeds Using Microsatellites. Genomics, 22(2):397-403.
Byme K. Chikhi L, Townsend SJ, Cruickshank RH. AI-derson GLH and Bruford MV (2004) Genetic diversity wit-hin and among European sheeps types and its
imp-Iications for breed conservalion, Molecular Ecology
(Basımda).
Comuet JM and Luikart G (1996) Description and power analysis of two tests for delecting recent population
bott-lenecks from allele frequency data, Genelics, 144(4),
2001-2014.
DIE (2002) TarımIstatistikleri Özeti, 1982-2001. DIE,
An-kara.
Diez-Tascon C, Littlejohn RP, Almeida PAR, Crawford AM (2000) Genetic vartatton wilhin lhe Merino sheep
breed:analysis of closely related populations using
mic-rosatellites. Anim Genet; 31, 243-251.
Farid A,O'Reilly E, Kelsey Jr. CR (2000) Genelric
analy-sis of ten sheep breeds using microsatellite markers.
Ca-nadian Journal of AnimalScience. 80. 9-17.
Goldslein DB and Schlölterer C (1998) Microsatellites:
Evolution and Application, Oxford University Press,
Ox-ford and Vienna.
Grigaliunaite i,Tapio M.Viinalass H. Grislis Z, Kantanen
J,Miceikienei(2003) Microsalellite varialion in lhe Baltic
sheep breeds. Veıerinarijair Zootechnlka, 21(43),66-73.
Gutierrez- Espeleta GA. Kalinowski ST, Boyce WM and
Hedrick PW (2000) Genetic variation and populalion struclure in desert bighorn sheep: implication for
con-servalion.Conservation Genetics, 1:3-15.
Li MH, Zhao SH, Bian C, Wang HS, Wei H, Liu B, Yu M,
Fan B,Chen SL,Zhu MJ, Li SJ. Xiong TA,Li K (2002)
Genetic relationships among twelve Chinese indigenous
goal populalions based on microsatellile analysis,Genet.
SeL. Evol., 34, 729-744.
Loftus RT,ErtugrulO, Harba AH,El-Barody MAA, Mac
-Hugh DE, Park SDE and Bradley DG (1999) A
rnic-rosatellile survey of cattle from a cenlre of origin:The
Near East. MolecularEcology, 8. 2015-2022.
Maudet C. Luikart G and Taberlet P (2002) Genelic d
i-versity and assignmenl test among seven French cattle
breeds based on microsalellite DNA analysis1, J. Anim. ScL. 80, 942-950.
Moazami-Goudarzi K, Laloe D, Furet JP, Grosclaude F (1997) Analysis of genetic relalionships between 10 caltle
Hasmer,lIasak ve Alma nSiyah Bl''' Koyun...
Nei M (1987) MolecularEvolutionary Genetes.Columbia University Press.Newyor1<.
Saitbekova N,Gaillard C. Obexer-Rufl G, Dolf G (1999)
Genetic diversity in Swiss goat breeds based on
mc-reseteaneanalysis,AnimGenel,30(1):36-4 1.Saitou N and NetM(1987)The neighbor-joiningmethod:a
new method for reconstnıction phylogenetic trees.
Mo-lecutar Biologyand Evoluti on,4:406-425.
SoysalMl.Togan
i.
Nizamlıoğlu M,Erqüven A. A1tunokV,Tuna YT, Özkan E, eürcan K, Bulut Z, Koban E (2001)
Türkiye Yerli veMelez Koyun IrklarınınGenetik Yapılarının
MikrosatellillerleIncelenm esi.VHAG-1553,TÜBITAK.
Tekin ME, Gürkan M, Karabulut O ve DuzgOn H (2001)
Merinos. Akkaraman ve lvesı Irkıarının Bazı Etçi Irklar ile
91
Melezierinde (Hasmer, Hasak, Hasıv ve linmer) Per
-formans Testve SeleksiyonÇalışmaları. ı. 001Verimi ve
YaşamaGOcü,Hayvancılık AraştırmaDergisi,2(1).1-8.
Togan I,SoysalMJ.Attunok V,Özkan E,Koban E(2003)
Populasyon genetiği çalışmalarında kullanılan yeni is·
tetlstik yöntemler,GAP3.Tarim Kongres i,2-3 Ekim, Har
-ranÜniversitesi Ziraat Fakültesi,Şanlıurfa.
YalçınBC (1986)Sheep and Goats in Turkey,Food and
Agriculture Organization of the United Nations, gom
e-Italy.
Yalç ın BC (1990) Koyun frklan" Koyun-Keçi Hastalıkları
ve Yetiştiriciliği", Aytuğ CN,Alaçam E, Özkoç Ü. Yalçın
BC, Gökçen H ve Türker H,TÜM VET Hayvancılık Hiz
-metleri Yayını, No:2, 387-422, Teknografik Matbaası, is