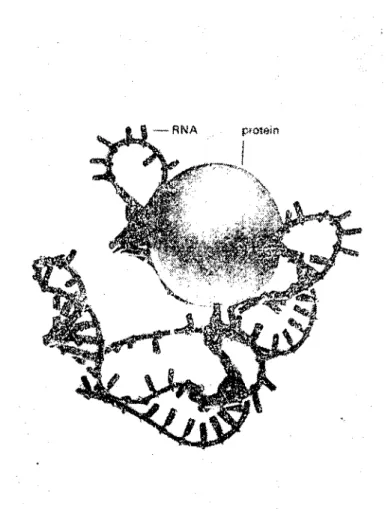
Başlık: RİBOSOMLARIN YAPISI VE GÖREVİYazar(lar):İSTANBULLUOĞLU, ErsinCilt: 23 Sayı: 1.2 DOI: 10.1501/Vetfak_0000001293 Yayın Tarihi: 1976 PDF
Tam metin
Şekil

Benzer Belgeler
Bir hükmi şahsın, buna rağmen, bir tek devletin hukuku ile diğer devletle- rinkiyle olduğundan daha sıkı bir şekilde bağlı olmayacak surette inşa edilmesi lâzımgeliyorsa,
Fakat tasarruf edilen gelir nisbeti artmış olsun veya olmasın, biz, herhangi modem bir cemiyetin temel psikolojik kuralı olarak ka bul ediyoruz ki, reel geliri arttığı zaman
1933 den sonra Almanya'da iştira kuvveti yaratmak suretiyle yapılan finansman o zamanki şartlar altında mal ve hizmet arzım kısa bir zamanda artırdığından, nominal milli
İthal mallarına olan talepteki azalma, yurtiçi har- camalardaki azalmanın kıymeti ile marjinal ithal temayülünün çar pımına (— A G. Deflasyon, yurtiçi fiyat seviye
Canlı ağırlık ve baş ölçüleri için en küçük kareler ortalamaları Tablo 1, vücut ölçülerine ait en küçük.. kareler ortalamaları Tablo
on the effects of vegetation elimination by grass carp (Ctenopharyngodon idella Val., 1844) on water quality, zooplankton and benthos. Kırkağaç M.U, Yavuzcan H.Y (2002): A
In light microscopic investigation, pecten was consisted of large number of capillaries and pigment cells (melanocytes) and the basal edge of pecten lined from the ventral part of
The purpose of this study is to reveal the effect of some acidifier agents as diluent solutions (therapeutic dose of ascorbic acid in distillated water and pure vinegar of
