FEN BĠLĠMLERĠ ENSTĠTÜSÜ
EKMEKLĠK BUĞDAYDA (Triticum aestivum L.) ÇĠMLENME
VE ERKEN GELĠġĠM DÖNEMĠNDE KURAKLIK VE TUZ
STRESĠNE TEPKĠLERĠN BELĠRLENMESĠ
Ece ĠYEM
YÜKSEK LĠSANS TEZĠ
TARLA BĠTKĠLERĠ ANABĠLĠM DALI
DĠYARBAKIR Haziran – 2019
I
TEġEKKÜR
Yüksek lisans eğitimime baĢlamamı sağlayan; bilgi ve önerilerini, desteğini esirgemeyen kıymetli danıĢman hocam Prof. Dr. Mehmet YILDIRIM’a teĢekkürlerimi sunarım.
AraĢtırmamın baĢından son aĢamasına kadar her an tecrübelerini, akademik biri-kimini paylaĢan, yardımlarını esirgemeyen, desteğini her zaman hissettiğim, çalıĢma prensibiyle yoluma ıĢık tutan değerli hocam Dr. Öğr. Üyesi Ferhat KIZILGEÇĠ’ye min-net ve Ģükranlarımı sunarım.
Çukurova Üniversitesi Bahçe Bitkilerimi Bölümü laboratuvarında yaptığım ça-lıĢmalarda bilgi ve tecrübelerini esirgemeyen Prof. Dr. Ebru KAFKAS hocama teĢek-kürlerimi sunarım.
ÇalıĢmalarımın her aĢamasında maddi ve manevi desteğini eksik etmeyen, labo-ratuvar çalıĢmalarında sürekli yanımda olup yardım eden değerli meslektaĢım ve mesai arkadaĢım Ragıp KÜÇÜK’e, Adana’da bulunduğum süre içerisinde her an yanımda olan meslektaĢım Tuğçe BAL ÖZTÜRK ve kıymetli ailesine, ihtiyacım olduğunda ya-nımda olan değerli mesai arkadaĢım Aykut DURMUġ’a ve laboratuvar çalıĢmalarımda yardımlarını eksik etmeyen Öğretim Görevlisi Nihan TAZEBAY ASAN’a teĢekkür ederim.
Her koĢulda, her zorlukta, enerjim tükendiğinde, motivasyonumu kaybettiğimde varlıkları ve güzel sözleri ile her zaman maddi, manevi yanımda olan canım anneme, babama ve kardeĢim Bahar ĠYEM'e tüm kalbimle teĢekkür ederim.
Adını sayamadığım fakat az ya da çok bir Ģekilde desteğini hissettiğim dostları-ma ve tüm sevdiklerime ayrıca teĢekkürlerimi sunarım.
II ĠÇĠNDEKĠLER TEġEKKÜR ... I ĠÇĠNDEKĠLER ... II ÖZET ……….IV ABSTRACT……….………V ÇĠZELGE LĠSTESĠ ... VI ġEKĠL LĠSTESĠ………..………...XIII KISALTMA VE SĠMGELER ... …….XVI
1. GĠRĠġ ... 1 1.1. Kuraklık Stresi ... 2 1.2. Tuz Stresi ... 4 2. KAYNAK ÖZETLERĠ ... 7 2.1. Kuraklık Stresi ... 7 2.2. Tuz Stresi ... 10 3. MATERYAL VE METOT ... 15 3.1. Materyal ... 15 3.1.1. Kimyasal Maddeler ... 15 3.1.2. Bitki Materyalleri ... 15 3.2. Metot ... 15
3.2.1. Bitki Materyalinin YetiĢtirilme Ortamı ... 15
3.2.2. Tuz Stresi Uygulamaları ... 17
3.2.3. Kuraklık Stresi Uygulamaları ... 17
3.2.4. Ġncelenen Özellikler ve Belirlenmesi ... 18
3.2.5. Verilerin Değerlendirilmesi ... 22
4. BULGULAR VE TARTIġMA ... 23
4.1. Kuraklık Stresinde Ġncelenen Özellikler ... 23
4.1.1. Kök Uzunluğu ... 23 4.1.2. Çim Kını Uzunluğu ... 25 4.1.3. Sürgün Uzunluğu ... 27 4.1.4. Kök YaĢ Ağırlığı ... 29 4.1.5. Sürgün YaĢ Ağırlığı ... 31 4.1.6. Turgor Ağırlığı ... 33 4.1.7. Kök Kuru Ağırlığı ... 34 4.1.8. Sürgün Kuru Ağırlığı ... 36 4.1.9. Oransal Su Ġçeriği (RWC) ... 38
III 4.1.10. Çimlenme Hızı ... 39 4.1.11. Çimlenme Gücü ... 41 4.1.12. Kök/Sürgün Uzunluk Oranı ... 43 4.1.13. Prolin Miktarı ... 45 4.1.14. Klorofil-a Miktarı ... 46 4.1.15. Klorofil-b Miktarı ... 48 4.1.16. Karotenoid Miktarı ... 50
4.1.17. Toplam Klorofil Miktarı ... 51
4.2. Tuz Stresinde Ġncelenen Özellikler ... 53
4.2.1. Kök Uzunluğu ... 53 4.2.2. Çim Kını Uzunluğu ... 55 4.2.3. Sürgün Uzunluğu ... 57 4.2.4. Kök YaĢ Ağırlığı ... 59 4.2.5. Sürgün YaĢ Ağırlığı ... 60 4.2.6. Turgor Ağırlığı ... 62 4.2.7. Kök Kuru Ağırlığı ... 64 4.2.8. Sürgün Kuru Ağırlığı ... 65 4.2.9. Oransal Su Ġçeriği (RWC) ... 67 4.2.10. Çimlenme Hızı ... 69 4.2.11. Çimlenme Gücü ... 70 4.2.12. Kök/Sürgün Uzunluk Oranı ... 72 4.2.13. Prolin Miktarı ... 73 4.2.14. Klorofil-a Miktarı ... 75 4.2.15. Klorofil-b Miktarı ... 77 4.2.16. Karotenoid Miktarı ... 78
4.2.17. Toplam Klorofil Miktarı ... 80
5. SONUÇ VE ÖNERĠLER ... 83
6. KAYNAKLAR ... 85
IV
ÖZET
EKMEKLIK BUĞDAYDA (Triticum aestivum L.) ÇĠMLENME VE ERKEN GELĠġĠM DÖ-NEMĠNDE KURAKLIK VE TUZ STRESĠNE TEPKĠLERĠN BELĠRLENMESĠ
YÜKSEK LĠSANS TEZĠ Ece ĠYEM DĠCLE ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ TARLA BĠTKĠLERĠ ANABĠLĠM DALI
2019
Günümüzde iklim değiĢimi dünya genelinde buğday üretimini olumsuz yönde etkile-mektedir. Bu olumsuzlukların baĢında kuraklık ve tuzluluk gibi abiyotik stres faktörleri ön plana çıkmaktadır. Gelecekte bu stres faktörlerine karĢı dayanıklı çeĢitlerin belirlenmesi ve geliĢtiril-mesi büyük önem arz etmektedir.
Bu çalıĢma, 5 ekmeklik buğday genotipinin (Lok-1, Lucilla, RSP-561, Envoy ve Wafia) çimlenme ve erken geliĢim döneminde kuraklık ve tuz stresine tepkilerinin belirlenmesi amacıy-la; 2018 yılında Çukurova Üniversitesi Ziraat Fakültesi Bahçe Bitkileri Bölümü laboratuarında tesadüf parselleri faktöriyel deneme desenine göre 3 tekrarlamalı olarak yürütülmüĢtür. Dene-mede, ekmeklik buğday genotiplerine 4 farklı kuraklık stresi (kontrol, -0.2, -0.4, -0.6 MPa, PEG-6000 çözeltisi kullanılarak) ve 4 farklı tuzluluk stresi (kontrol, 4, 8 ve 12 dS/m, NaCl çö-zeltisi kullanılarak) uygulanmıĢtır.
AraĢtırmada çim kını uzunluğu, kök uzunluğu, sürgün uzunluğu, kök yaĢ ve kuru ağırlı-ğı, sürgün yaĢ ve kuru ağırlıağırlı-ğı, çimlenme hızı, çimlenme gücü, oransal nem içeriği, turgor ağır-lığı, klorofil-a, klorofil-b, karotenoid ve toplam klorofil içeriklerinde ve prolin içeriği özellikleri incelenmiĢtir.
ÇalıĢmadan elde edilen verilere göre tuz ve kuraklık stresinde kök/sürgün oranı özelliği hariç diğer incelenen özelliklerin tümünde istatistiksel olarak önemli farklılıklar görülmüĢtür. Kuraklık ve tuz stresi artıĢına bağlı olarak, kök uzunluğu, sürgün uzunluğu, kök yaĢ ve kuru ağırlığı, sürgün yaĢ ve kuru ağırlığı, çimlenme hızı, çimlenme gücü, oransal nem içeriği, turgor ağırlığı, klorofil-a,klorofil-b, karotenoid ve toplam klorofil içeriklerinde kontrol koĢulları ile karĢılaĢtırıldığında incelene bu özelliklerde azalma meydana geldiği görülmüĢtür. Çim kını uzunluğu ve prolin içeriği özelliğinde ise artıĢ meydana gelmiĢtir. Genotipler arasında kuraklık ve tuzluluk stresine en iyi yanıt veren genotip RSP-561 olduğu, klorofil-a, klorofil-b, karotenoid ve toplam klorofil miktarında ise Lok-1 çeĢidi olumlu tepki verdiği görülmüĢtür. Lucilla çeĢidi ise, kuraklık ve tuzluluğa hassas olduğu belirlenmiĢtir.
Yapılan araĢtırma sonucunda kuraklık ve tuzluluk dozu artıĢı ekmeklik buğday genotip-lerinin erken geliĢim dönemlerinde olumsuz yönde etkilediği, stres koĢulları artıĢına bağlı ola-rak dirençli genotiplerde prolin miktarında artıĢ meydana getirmiĢtir.
V
ABSTRACT
DETERMINATION OF RESPONSE TO DROUGHT AND SALT STRESS ON GERMINATION AND EARLY GROWTH STAGE IN BREAD WHEAT (Triticum
aestivum L.)
MASTER’S THESIS Ece IYEM
DEPARTMENT OF FIELD CROPS
INSTITUTE OF NATURAL AND APPLIED SCIENCES UNIVERSITY OF DICLE
2019
In recently, climate change has a negative impact on wheat production worldwide. Abi-otic stress factors such as drought and salinity come to the forefront. It is of great importance to identify and develop varieties resistant to these stress factors in the future.
In this study, in order to determine the response of drought and salt stress in 5 bread wheat genotypes (Lok-1, Lucilla, RSP-561, Envoy and Wafia) during germination and early growth stage; was carried out according to completely randomized factorial design with three replications, at Cukurova University, Faculty of Agriculture, Department of Horticulture in 2018. In the experiment, 4 different drought stresses (control, -0.2, -0.4, -0.6 MPa, by using PEG-6000 solution) and 4 different salinity stress (control, 4, 8 and 12 dS / m by using NaCl solution) was applied to bread wheat genotypes.
In the research, root length, coleoptile length, shoot length, root fresh and dry weight, shoot fresh and dry weight, germination rate, seed vigor, relative water content, turgor weight, chlorophyll-a, chlorophyll-b, carotenoid and total chlorophyll content and proline content were examined.
According to the data obtained from the study, statistically significant differences were observed in all other examined properties, except the root / shoot rate feature in salt and drought stress. Depending on increase in drought and salt stress, root length, shoot length, root fresh and dry weight, shoot fresh and dry weight, germination rate, seed vigor, relative water content, turgor weight, chlorophyll-a, chlorophyll-b, carotenoid and total chlorophyll content was com-pared to control conditions, it was observed that these properties decreased. In case, coleoptile length and proline content was increased. Among genotypes, the genotype RSP-561 was the best response to drought and salinity stress, whereas chlorophyll-a, chlorophyll-b, carotenoid and total chlorophyll were found to be positive in the Lok-1 variant. The Lucilla cultivar was found to be sensitive to drought and salinity.
As a result of the research, increase in drought and salinity dose negatively affected the bread wheat genotypes in germination and early growth stage, was caused an increase in the proline amount in resistant genotypes due to increased stress conditions.
VI
ÇĠZELGE LĠSTESĠ
Çizelge No Sayfa
Çizelge 3.1. Denemede Kullanılan Ekmeklik Buğday Genotipleri 15
Çizelge 3.2. Denemede Uygulanan NaCl Çözeltileri 17
Çizelge 3.3. Denemede Uygulanan PEG Çözeltileri 17
Çizelge 4.1. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Kök Uzunluğuna Ait
Varyans Analiz Sonuçları 23
Çizelge 4.2. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Kök Uzunluklarına (cm) Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 24 Çizelge 4.3. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik
Buğday Genotiplerinin Çimlenme Dönemindeki Çim Kını Uzunluğuna Ait
Varyans Analiz Sonuçları 25
Çizelge 4.4. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Çim Kını Uzunluklarına (cm) Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 26 Çizelge 4.5. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik
Buğday Genotiplerinin Çimlenme Dönemindeki Sürgün Uzunluğuna Ait
Varyans Analiz Sonuçları 27
Çizelge 4.6. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Sürgün Uzunluklarına (cm) Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 28 Çizelge 4.7. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik
Buğday Genotiplerinin Çimlenme Dönemindeki Kök YaĢ Ağırlığın Ait
Varyans Analiz Sonuçları 29
Çizelge 4.8. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Kök YaĢ Ağırlığına (g) Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 30 Çizelge 4.9. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik
Buğday Genotiplerinin Çimlenme Dönemindeki Sürgün YaĢ
AğırlığınaAitVaryans Analiz Sonuçları 31
Çizelge 4.10. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Sürgün YaĢ Ağırlığına (g) Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 32
VII
Çizelge 4.11. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Turgor Ağırlığına Ait
Varyans Analiz Sonuçları 33
Çizelge 4.12. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Turgor Ağırlığına (g) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 33
Çizelge 4.13. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Kök Kuru Ağırlığına Ait
Varyans Analiz Sonuçları 34
Çizelge 4.14. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Kök Kuru Ağırlığına (g) Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 35 Çizelge 4.15. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik
Buğday Genotiplerinin Çimlenme Dönemindeki Sürgün Kuru Ağırlığına Ait
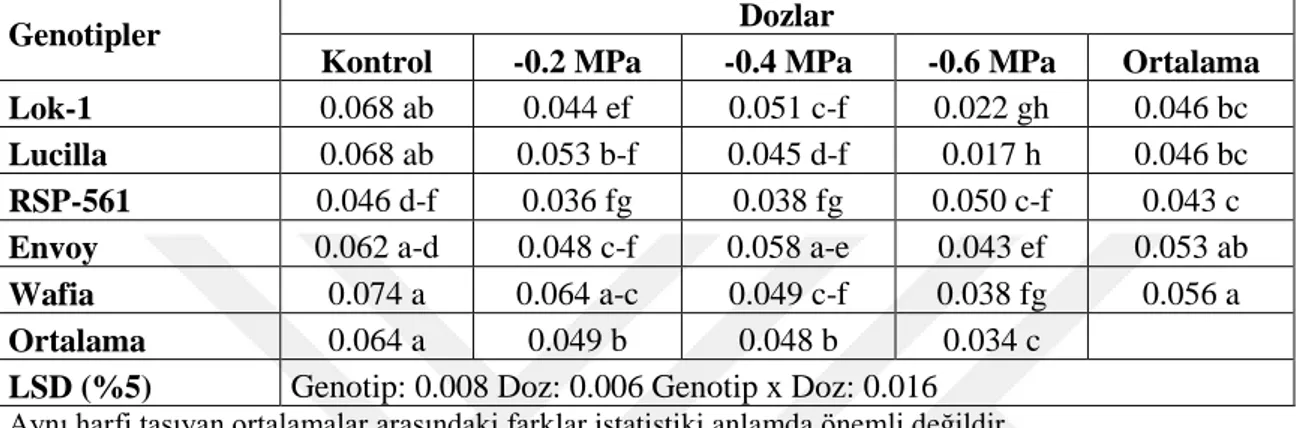
Varyans Analiz Sonuçları 36
Çizelge 4.16. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Sürgün Kuru Ağırlığına (g) Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 37 Çizelge 4.17. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik
Buğday Genotiplerinin Çimlenme Dönemindeki Oransal Su Ġçeriğine Ait
Varyans Analiz Sonuçları 38
Çizelge 4.18. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Oransal Su Ġçeriğine Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 39
Çizelge 4.19. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Çimlenme Hızına Ait
Varyans Analiz Sonuçları 40
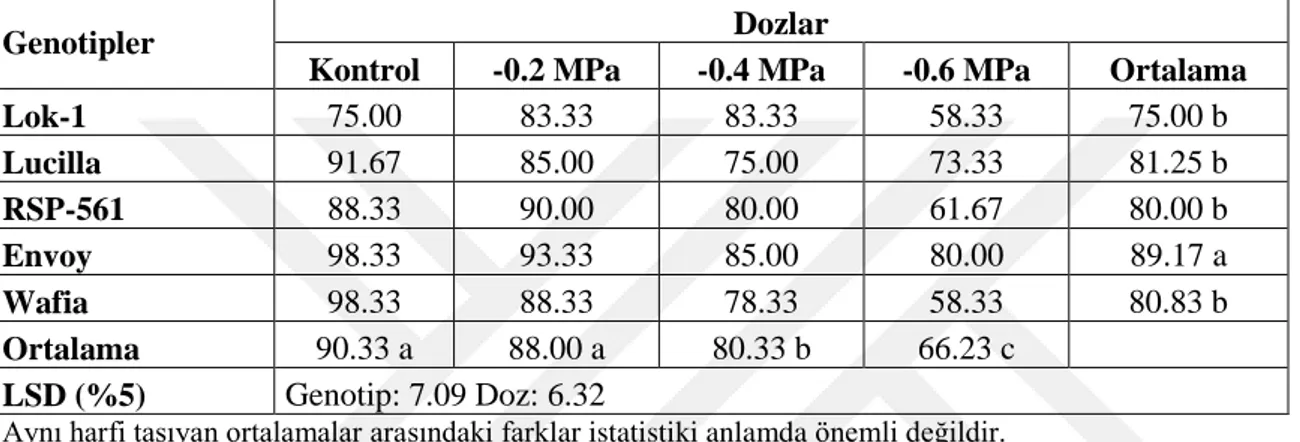
Çizelge 4.20. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemdeki Çimlenme Hızına (%) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 40
Çizelge 4.21. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Çimlenme Gücüne
VIII
Çizelge 4.22. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemdeki Çimlenme Gücüne (%) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 42
Çizelge 4.23. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Kök/Sürgün Uzunluk
Oranına Ait Varyans Analiz Sonuçları 43
Çizelge 4.24. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemdeki Kök/Sürgün Uzunluk Oranına Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 44 Çizelge 4.25. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik
Buğday Genotiplerinin Çimlenme Dönemindeki Prolin Miktarına Ait
Varyans Analiz Sonuçları 45
Çizelge 4.26. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemdeki Prolin Miktarına (µmol/g)Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 45 Çizelge 4.27. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik
Buğday Genotiplerinin Çimlenme Dönemindeki Klorofil-a Miktarına Ait
Varyans Analiz Sonuçları 46
Çizelge 4.28. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemdeki Klorofil-a Miktarına Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 47
Çizelge 4.29. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Klorofil-b Miktarına Ait
Varyans Analiz Sonuçları 48
Çizelge 4.30. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemdeki Klorofil-b Miktarına Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 49
Çizelge 4.31. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Karotenoid Miktarına Ait
Varyans Analiz Sonuçları 50
Çizelge 4.32. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemdeki Karotenoid Miktarına Ait Ortalama
IX
Çizelge 4.33. Farklı PEG-6000 Dozu Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemindeki Toplam Klorofil Miktarına
Ait Varyans Analiz Sonuçları 51
Çizelge 4.34. Farklı Kuraklık Uygulamalarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemdeki Toplam Klorofil Miktarına Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 52 Çizelge 4.35. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday
Genotiplerinin Çimlenme Dönemi Kök Uzunluğuna Ait Varyans Analiz
Sonuçları 53
Çizelge 4.36. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Kök Uzunluğuna (cm) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 54
Çizelge 4.37. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Çim Kını Uzunluğuna Ait Varyans Analiz
Sonuçları 55
Çizelge 4.38. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Çim Kını Uzunluğuna (cm) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 56
Çizelge 4.39. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Sürgün Uzunluğuna Ait Varyans Analiz
Sonuçları 57
Çizelge 4.40. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Sürgün Uzunluğuna (cm) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 58
Çizelge 4.41. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Kök YaĢ Ağırlığına Ait Varyans Analiz
Sonuçları 59
Çizelge 4.42. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Kök YaĢ Ağırlığına (g) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 59
Çizelge 4.43. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Sürgün YaĢ Ağırlığına Ait Varyans
X
Çizelge 4.44. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Kök Uzunluğuna (cm) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 61
Çizelge 4.45. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Turgor Ağırlıklarına Ait Varyans Analiz
Sonuçları 62
Çizelge 4.46. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Turgor Ağırlıklarına (g) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 63
Çizelge 4.47. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Kök Kuru Ağırlığına Ait Varyans Analiz
Sonuçları 64
Çizelge 4.48. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Kök Kuru Ağırlığına (g) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 64
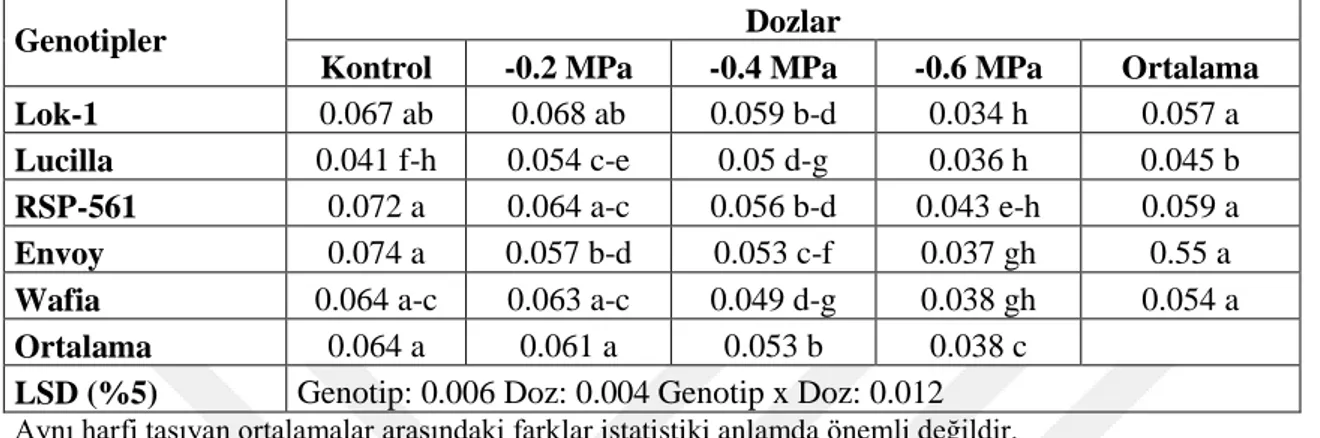
Çizelge 4.49. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Sürgün Kuru Ağırlığına Ait Varyans
Analiz Sonuçları 66
Çizelge 4.50. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Sürgün Kuru Ağırlığına (g) Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 66 Çizelge 4.51. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday
Genotiplerinin Çimlenme Dönemi Oransal Su Ġçeriğine Ait Varyans Analiz
Sonuçları 67
Çizelge 4.52. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Oransal Su Ġçeriğine Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 68
Çizelge 4.53. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Çimlenme Hızına Ait Varyans Analiz
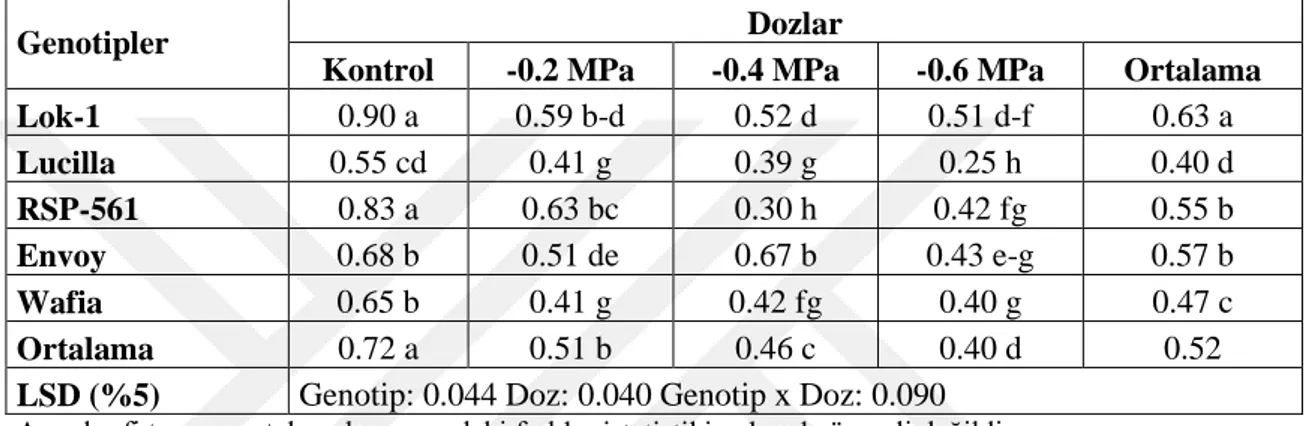
Sonuçları 69
Çizelge 4.54. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Çimlenme Hızına (%) Ait Ortalama
XI
Çizelge 4.55. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Çimlenme Gücüne Ait Varyans Analiz
Sonuçları 71
Çizelge 4.56. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Çimlenme Gücüne (%) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 71
Çizelge 4.57. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Kök/Sürgün Uzunluk Oranına Ait
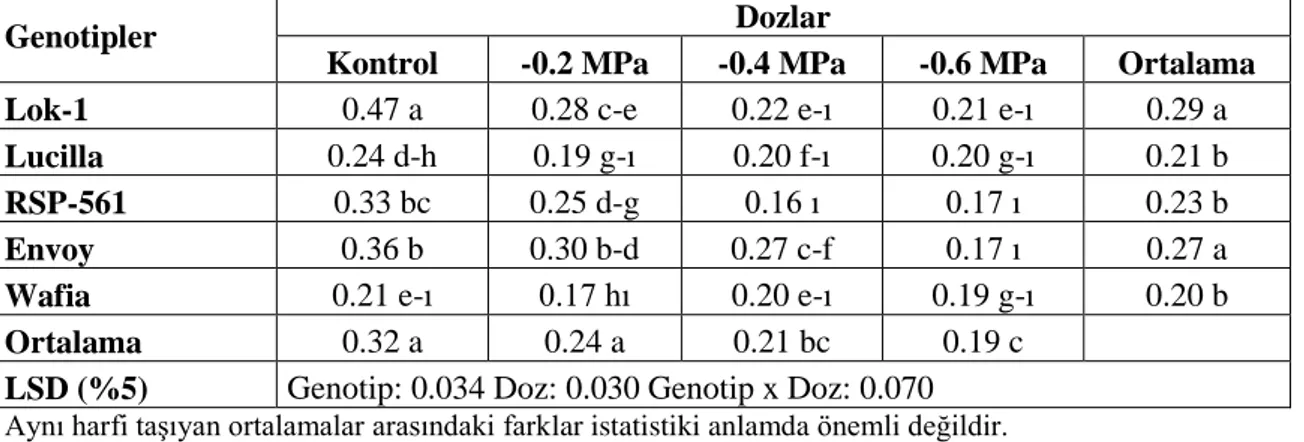
Varyans Analiz Sonuçları 72
Çizelge 4.58. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Kök/Sürgün Uzunluk Oranına Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 73 Çizelge 4.59. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday
Genotiplerinin Çimlenme Dönemi Prolin Miktarına Ait Varyans Analiz
Sonuçları 74
Çizelge 4.60. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Prolin Miktarına (µmol/g) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 74
Çizelge 4.61. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Klorofil-a Miktarına Ait Varyans Analiz
Sonuçları 75
Çizelge 4.62. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Klorofil-a Miktarına (mg/g) Ait Ortalama
Değerler ve Çoklu KarĢılaĢtırmalar 76
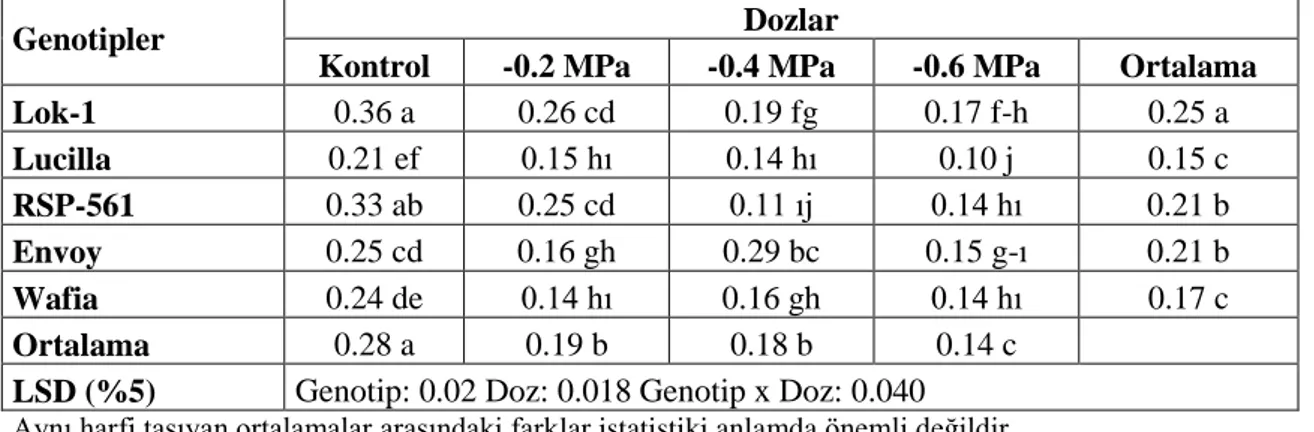
Çizelge 4.63. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Klorofil-b Miktarına Ait Varyans Analiz
Sonuçları 77
Çizelge 4.64. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Klorofil-b Miktarına (mg/g) Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 77 Çizelge 4.65. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday
Genotiplerinin Çimlenme Dönemi Karotenoid Miktarına Ait Varyans Analiz
Sonuçları 78
Çizelge 4.66. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Karotenoid Miktarına (mg/g) Ait
XII
Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 79 Çizelge 4.67. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday
Genotiplerinin Çimlenme Dönemi Toplam Klorofil Miktarına Ait Varyans
Analiz Sonuçları 80
Çizelge 4.68. Farklı Tuz Konsantrasyonlarında YetiĢtirilen Ekmeklik Buğday Genotiplerinin Çimlenme Dönemi Toplam Klorofil Miktarına (mg/g) Ait Ortalama Değerler ve Çoklu KarĢılaĢtırmalar 81
XIII
ġEKĠL LĠSTESĠ
ġekil No Sayfa
ġekil 3.1. Dezenfekte edilmiĢ ekmeklik buğday tohumlarının cam petrilere
yerleĢtirilmesi 16
ġekil 3.2. Ġklim kabininde çimlendirmeye bırakılan ekmeklik buğday tohumları 16 ġekil 3.3. Cam petrilere yerleĢtirilen tohumlara PEG çözeltisi uygulaması 18 ġekil 3.4. Denemenin 8. gününde çimlenen buğdayların kumpas yardımı ile
ölçümlerinin yapılması 19
ġekil 3.5. Çimlendirme denemesinde her gün gözlem alınması 20 ġekil 3.6. Pigment içeriğinin belirlemek için bitki materyali ekstraksiyon iĢlemi 21 ġekil 3.7. Prolin içeriğinin belirlenmesinde yapılan iĢlemler 22 ġekil 4.1. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin kök
uzunluklarında meydana gelen değiĢimler 25
ġekil 4.2. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin çim kını
uzunluklarında meydana gelen değiĢimler 27
ġekil 4.3. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin sürgün
uzunluklarında meydana gelen değiĢimler 29
ġekil 4.4. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin kök yaĢ
ağırlıklarında meydana gelen değiĢimler 31
ġekil 4.5. Kuraklık stresine maruz ekmeklik buğday genotiplerinin sürgün yaĢ
ağırlıklarında meydana gelen değiĢimler 32
ġekil 4.6. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin turgor
ağırlıklarında meydana gelen değiĢimler 34
ġekil 4.7. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin kök kuru
ağırlıklarında meydana gelen değiĢimler 36
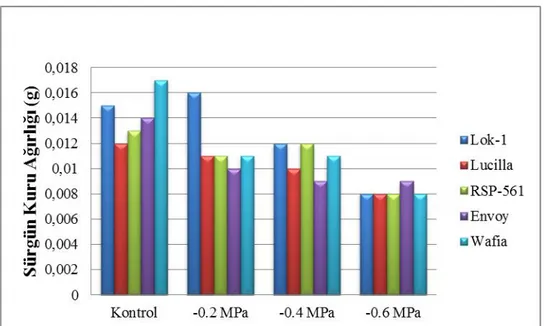
ġekil 4.8. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin sürgün kuru
ağırlıklarında meydana gelen değiĢimler 38
ġekil 4.9. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin oransal su
içeriklerinde meydana gelen değiĢimler 39
ġekil 4.10. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerininçimlenme
hızlarında meydana gelen değiĢimler 41
ġekil 4.11. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin çimlenme
gücünde meydana gelen değiĢimler 43
ġekil 4.12. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin kök/sürgün uzunluk oranlarında meydana gelen değiĢimler 44
XIV
ġekil 4.13. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin prolin
miktarlarında meydana gelen değiĢimler 46
ġekil 4.14. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin klorofil-a
miktarlarında meydana gelen değiĢimler 48
ġekil 4.14. Kuraklık stresi altındaki ekmeklik buğday genotiplerinin klorofil-b miktarlarında meydana gelen değiĢimler 49 ġekil 4.16. Kuraklık stresine maruz kalan buğday genotiplerinin karotenoid
miktarlarında meydana gelen değiĢimler 51 ġekil 4.17. Kuraklık stresine maruz kalan ekmeklik buğday genotiplerinin toplam
klorofil miktarlarında meydana gelen değiĢimler 53 ġekil 4.18. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin kök
uzunluklarında meydana gelen değiĢimler 55
ġekil 4.19. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin çim kını
uzunluklarında meydana gelen değiĢimler 57
ġekil 4.20. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin sürgün
uzunluklarında meydana gelen değiĢimler 58
ġekil 4.21. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin kök yaĢ
ağırlıklarında meydana gelen değiĢimler 60
ġekil 4.22. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin sürgün yaĢ
ağırlıklarında meydana gelen değiĢimler 62
ġekil 4.23. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin turgor
ağırlıklarında meydana gelen değiĢimler 63
ġekil 4.24. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin kök kuru
ağırlıklarında meydana gelen değiĢimler 65
ġekil 4.25. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin sürgün kuru ağırlıklarında meydana gelen değiĢimler 67 ġekil 4.26. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin oransal su
içeriği ağırlıklarında meydana gelen değiĢimler 68 ġekil 4.27. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin çimlenme
hızlarına meydana gelen değiĢimler 70
ġekil 4.28. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin çimlenme
gücünde meydana gelen değiĢimler 72
ġekil 4.29. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin kök/sürgün uzunluk oranlarında meydana gelen değiĢimler 73
XV
ġekil 4.30. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin prolin
miktararında meydana gelen değiĢimler 75
ġekil 4.31. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin klorofil-a
miktarlarında meydana gelen değiĢimler 76
ġekil 4.32. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin klorofil-b
miktarlarında meydana gelen değiĢimler 78
ġekil 4.33. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin karotenoid
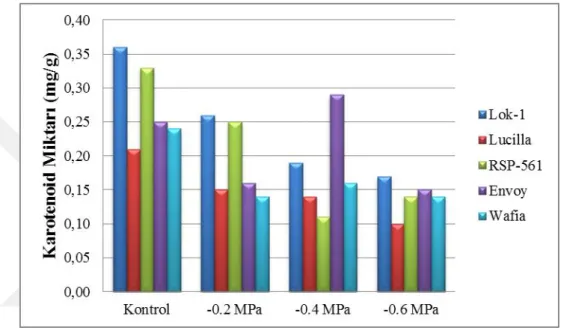
miktarlarında meydana gelen değiĢimler 80
ġekil 4.34. Artan tuz stresine maruz kalan ekmeklik buğday genotiplerinin toplam klorofil miktarlarında meydana gelen değiĢimler 81
XVI KISALTMA VE SĠMGELER cm : Santimetre g : Gram mg : Miligram K : Potasyum Na : Sodyum da : Dekar
TÜĠK : Türkiye Ġstatistik Kurumu
FAO : Food and Agriculture Organization ºC : Santigrad derece ml : Mililitre µmol : Mikromol µl : Mikrolitre dS : Desisimens m : Metre MPa : Megapascal
pH : Toprağın asitlik-bazlık derecesi L : Litre
A : Absorbans değer,
1
1. GĠRĠġ
Buğday iyi bir besin kaynağı olması, adaptasyonunun geniĢliği, üretiminin kolay olması gibi nedenlerle hem dünya hem de ülkemiz tarımında önemli bir yere sahiptir (Anonim, 2017). Ayrıca, dünyada ve ülkemizde tarımsal sanayide hammadde olarak kullanılması, kırsal bölgelerin en önemli gelir kaynağı olması bakımından stratejik öneme sahip tarımsal ürün konumundadır (TaĢçı ve ark., 2017).
Dünya üzerinde üretilen buğdayın % 95’ini hekzaploid ekmeklik buğday (2n = 6x = 42, BBAADD) oluĢturmakta ve bunu kültürü yapılan diğer buğday türlerden tetra-ploid makarnalık buğday (2n = 4x = 28, BBAA) takip etmektedir (Braun ve Sãulescu, 2002; Braun ve ark., 2010).
Dünyada ve ülkemiz tarımında önemli tahıl ürünlerinden olan buğday; ülkemizde 77 milyon da ekim alanında 21.5 milyon ton üretimi yapılmaktadır (TUĠK 2017). Ekmeklik buğday ise, ekiliĢ alanın % 84’ünü, üretiminin % 82’sini oluĢturmak-tadır (YakıĢır ve ark., 2016). Bu nedenle insan beslenmesinde geçmiĢten günümüze ka-dar vazgeçilmez bir gıda maddesi olan buğdayın, veriminde sağlanacak artıĢ ülkemiz ekonomisine önemli katkılar sağlayacaktır.
Uygun koĢullar altında geliĢimini en iyi biçimde gösteren bitkilerde, beklenmed-ik koĢullar oluĢtuğunda fizyolojbeklenmed-ik değiĢimler meydana gelir ve bu istenmeyen değiĢim-lere stres denir (Çulha ve Çakırlar, 2011). Buğday verimini sınırlayan stres etmenleri biyotik etmenler (mikroorganizmalar, mantar, böcek vb.) ve abiyotik etmenler (sıcaklık, su, tuz, ıĢık, pH gibi) olmak üzere iki gruba ayrılır (Kadıoğlu, 2011). Tarımda verimi ve ürün kaybını etkileyen abiyotik stres faktörleri arasında % 26 oranla en fazla kuraklık stresi görülmekte ve bunu da % 20’lik oranla tuz stresi takip etmektedir (Blum, 1985; Ashraf, 1994). Tarımsal amaçla kullanılan alanların yalnızca % 90’ı mutlaka bir stres faktöründen etkilenmektedir (Örs ve Ekinci, 2015).
2
1.1. Kuraklık Stresi
Kuraklık, abiyotik stres faktörleri arasında ürün verimliliğini en fazla etkileyen stres faktörü olarak kabul edilir (Lambers ve ark., 2008). Meteorolojik, tarımsal, sosyo-ekonomik ve hidrolojik olarak farklı tanımlanabilen kuraklık, optimum değerlerden da-ha düĢük olan yağıĢ miktarları sebebiyle su kaynaklarının ve arazilerin olumsuz olarak etkilenmesi gibi genel bir tanımı bulunmaktadır (Tuberosa, 2012; Turner ve ark., 2014). Tarımsal kuraklık ise bitkinin büyüme ve geliĢmesinde yaĢam döngüsünü tamamlaya-bilmesi gerekli olan nem miktarındaki eksiklik olarak tanımlanmaktadır (Manivannan ve ark., 2008).
Tuzluluk, kuraklık ve sıcaklık gibi abiyotik stresler bitkinin büyüme ve geliĢmesini sınırlayan ana faktörlerdendir. Verimi etkileyen birçok fizyolojik karakterin de kuraklık koĢullarından etkilendiği ve su sıkıntısı hassasiyetinin en fazla generatif dönemde etkili olduğu bilinmektedir. Yapılan araĢtırmalara göre tohum oluĢum döne-minde gerçekleĢen Ģiddetli kuraklığın ciddi verim kayıplarına yol açtığını, çiçeklenme döneminde meydana gelen su noksanlığının bitkilerde kısırlığa neden olduğunu ortaya koymaktadır (Farooq ve ark., 2009a). Kurak Ģartlarda bitki büyümesinin, bitkinin su sıkıntısını yaĢadığı süreye ve döneme bağlı olarak önemli ölçüde etkilenir. Ġlk dönem-lerde oluĢan kuraklıkta, bitki ihtiyacı olan suya daha çok ulaĢabilmek için kök geliĢimi-ni hızlandırır ve gövde uzamasını yavaĢlatır (Öztürk, 2015). Hasara yol açabilecek ka-dar uzun süren kurak koĢullar nedeniyle bitkide kök ve sürgün geliĢimi durur, yaprak sayısı ve alanı azalır, daha sonraki sürede sararma ve dökülme meydana gelir (Anjumve ark., 2011; Öztürk, 2015). Bitki büyümesindeki bu azalma hücre bölünmesinin ve geniĢlemesinin durmasına bağlı olarak doğrudan fotosentez oranında düĢmeye neden olur (Anjum ve ark., 2011).
Bitki kuraklık stresini algılamaya baĢladığında verdiği tepkilerden bir tanesi stomaların kapanması olayıdır (Osakabe ve ark., 2014). Kuraklık stresinden etkilenen bitkiler stomalarını kapatarak veya kısarak terleme ve su kaybını en aza indirmeye çalıĢmakla birlikte, bitki için fotosentezde gerekli olan karbondioksitin alımının azal-masına neden olmaktadır. Bu da fotosentez oranında düĢüĢe neden olur, büyüme ve geliĢmeyi olumsuz etkiler (Farooq ve ark., 2009a; Dolferus, 2014).
3
Bitkiler ortamdaki su eksikliğini algıladıklarında; dokulardaki su miktarını ve turgor basıncını azaltarak büyüme ve geliĢmeyi devam ettirmeye çalıĢır (Kiani ve ark. 2007; Hussain ve ark. 2009). Kuraklık ve tuzluluk stresi altında, mevcut su miktarını azaltmadan, düĢük su potansiyeline yardımcı olan organik ve inorganik çözücülerin birikimine osmotik ayarlama denir (Serraj ve Sinclair, 2002). Bununla birlikte, osmolit olarak adlandırılan bu çözücüler membranlar, enzimler ve diğer makromoleküller üzerinde, yüksek konsantrasyonda bile zararlı etkiler oluĢturmazlar ve bunlara uyumlu çözücüler de denir (Cechin ve ark., 2006; Kiani ve ark., 2007). Bu uyumlu çözeltiler arasında çözünür Ģekerler, Ģeker alkolleri, prolin, glisinbetain, organik asitler vb. bulu-nur (Cechin ve ark., 2006; Kiani ve ark., 2007; Farooq ve ark., 2008, 2009a,2009b). Bitkinin kuraklık stresine maruz kaldığı durumda su dengesini koruyan ozmolitler, doğrudan toleransı arttırmazlar. Yapraktaki su basıncını dengeleyerek stoma iletkenliğini arttırarak, fotosentezin devam etmesini sağlar ve böylece büyümenin de-vamlılığını sağlar.
Bu durum ozmolitlerin stres koĢullarında bitkilere dayanıklılık sağlamaktadır. Stres koĢullarının uzun süre devam ettiği durumlarda ise ozmolitler turgor basıncının kaybını dengelemekte yeterli olmamaktadır. Kuraklık stresi oluĢtuğu anda ilk olarak biriken amino asitlerden olan prolin, bir osmolit veya bir ozmoprotektan olarak sınıflandırılan birkaç küçük molekülden biridir (Csonka, 1989; Anjum ve ark., 2011; Liang ve ark., 2013; Kishor ve Sreenibasulu, 2014). Abiyotik stres faktörlerinin ortak bir cevabı olan prolin birikmesi aynı zamanda polen ve tohum geliĢimi sırasında yüksek oranda görüldüğü belirtilmiĢtir. Bu nedenle prolin birikiminin doğal geliĢim sürecinde mi ortaya çıktığı ya da stres koĢulları altında mı indüklendiği kesinleĢmemiĢtir. Fakat her iki durumda da prolin birikiminin düzenlenmesi, sentezindeki hız sınırlayıcı enzim olan P5CS'nin etkinliğine bağlıdır (Delauney ve Verma 1993; Szekely ve ark., 2008).
Bitkilerin kuraklık stresine olan tepkilerin karmaĢık olması diğer önemli abiyotik stres faktörlerine tepkilerinin benzer sinyal iletim ve mekanizmalarını kullanmalarından ileri gelir. Bunun sonucunda tarla koĢullarında kuraklık, tuzluluk ve sıcaklık streslerinin bitkiye aynı dönemlerde etki ettiği ve bu nedenle stres toleransının arttırılmasına yönelik çalıĢmalarda ayrı düĢünülemeyeceği göz ardı edilmemelidir (Suzuki ve ark., 2014).
4
1.2. Tuz Stresi
Çözünebilir tuz iyonlarının kurak ve yarı kurak bölgelerde yıkanarak yüksek taban suyu ile kapilarite yoluyla toprak yüzeyine çıkması ve daha sonra buharlaĢma sonucu suyun topraktan ayrılarak tuzun toprak yüzeyinde birikmesi olayına toprak tu-zluluğu denir (Ergene, 1982; Kantarcı, 2000; Ekmekçi ve ark., 2005). Tuzlu olarak sınıflandırılan topraklar; EC değerinin 4 dS / m ve daha fazla olduğu, yaklaĢık olarak 40 mM NaCl ve 0.2 MPa osmotik basınca eĢ değer topraklar olarak sınıflandırılmaktadır (Munns ve Tester, 2008). Dünyanın birçok yerinde buğdayda ciddi verim kayıplarına neden olan tuzluluk, abiyotik stresler arasında dünya çapında 800 milyon hektardan fazla alanı olumsuz etkiler ve tarımsal sürdürülebilirlik için büyük bir tehdittir (Munns ve Tester, 2008). Dünyadaki toplam arazi miktarının % 6.5'i ve sulu arazilerin % 19.5'i tuzdan etkilenmiĢtir (FAO, 2018). Bununla birlikte, Bouthour ve ark. (2015) ekilebilir alanların % 50’sinin, 2050 yılına kadar tuzluluktan etkileneceğini öngörmüĢlerdir.
Bitki büyüme ve geliĢmesinde önemli etkiye sahip olan tuz stresi; tuza maruz kalan bitkilerde tohum çimlenmesi, kök uzunluğu ve ağırlığının, sürgün uzunluğu ve ağırlığının azalmasına neden olur (Liang ve ark., 2014). Yüksek tuzluluk, bitkilerde; su stresi, iyon toksisitesi, beslenme bozuklukları, oksidatif stres, metabolik faaliyetlerin değiĢmesi, membran düzensizliği, hücre bölünmesi ve geliĢmesinin azalması gibi çeĢitli etkilere yol açar (Hasegawa ve ark., 2000; Munns, 2002; Zhu, 2007). Tuz stresine ma-ruz kalan bitkilerde bitki büyümesinin etkilendiği anda ilk olarak osmotik stres oluĢur (Horie ve ark., 2011). Kök rizosferinde tuz miktarının artmasıyla meydana gelen osmot-ik stres, kullanılabilir su içeriğini azaltır ve fizyolojosmot-ik kuraklığa neden olur (Tuteja, 2007). Osmotik stresin oluĢmasından sonra ortamda Na+
ve Cl- iyonları artar ve diğer besin elementleri ile rekabete girer. Bunun sonucunda bitkilerde iyon dengesizliği veya iyon noksanlığı ortaya çıkar (Hu ve Schmidhalter, 2005). Tuz stresi; bitkiler üzerindeki birincil etkisini iyon ve osmotik stres, ikincil etkisini ise bu stres etkenleri neticesinde ortaya çıkan toksik bileĢiklerin sentezlenmesi ve bir takım yapısal bozulmalar ile gösterir. Meydana gelen bu ikincil etkiler; klorofil, protein ve zar iĢleyiĢine zarar veren aktif oksijen türleri, K+
alımının engellenmesi, hücre ölümü, fotosentezin yavaĢlaması ve metabolik toksisitedir (Hong ve ark., 2009; Botella ve ark. 2005).
5
Tuz stresinin doğrudan etki ettiği bitkilerde, hücre büyümesi ve geniĢlemesinin engellenmesi sonucunda kök geliĢiminde azalma meydana gelir (Wang ve ark., 2009). Tuz stresinin bitki köklerine direkt etki etmesine rağmen, yaprak ve sürgün geliĢimi tuzluluğa karĢı daha hassastır. Bu durum tuz stresinde bitkilerdeki kök/sürgün oranının artmasına neden olur (Munns ve Tester, 2008). Bitki büyüme ve geliĢmesinde tohum çimlenmesi ve fide büyümesi en kritik iki aĢamadır (Hubbard ve ark., 2012). Bu önemli aĢamalar abiyotik stres faktörlerine en duyarlı olanlardır (Patade ve ark., 2011). Tuz stresi ortamdaki osmotik potansiyeli azaltarak, Na+ ve Cl- içeren iyonların artmasıyla bitki geliĢimi için gerekli K+
ve Ca+2 gibi iyonların alımı engellenmekte bu da tohum çimlenmesini etkilemektedir (Huang ve Redmann, 1995).
Bitkiler oluĢan tuz stresine karĢı moleküler ve biyokimyasal tolerans mekaniz-maları geliĢtirmiĢlerdir. Biyokimyasal tolerans mekanizmekaniz-maları; iyonların bitki bünyesinde veya belirli bölümlerde birikmesi, köklerden iyon alımı kontrolü, osmotik düzenleyiciler ile antioksidanlardan oluĢur (Parida ve Das, 2005).
7
2. KAYNAK ÖZETLERĠ 2.1. Kuraklık Stresi
Jajarmi (2009), Ġran’da yedi buğday çeĢidi (azar, Gv 3-20, dorom, omid, Keraceharvand, tabaci, arvand) ile yaptığı çalıĢmada kuraklık stresinin çimlenme in-deksleri üzerindeki etkilerini incelemiĢtir. Polietilen-glikol (PEG-6000) ile altı seviyede (damıtılmıĢ su, -3, -6, -9, -12, -15 bar) kuraklık uygulaması yapılmıĢ, tüm özelliklerde stres düzeyindeki artıĢ ile önemli bir düĢüĢ gözlendiği belirtilmiĢtir. Diğer özellikler arasında kök uzunluğunun kuraklık stresine daha duyarlı olduğu; çimlenme yüzdesi incelendiğinde ise, -3 bar seviyesinde kuraklık stresine yüksek tolerans gösterdiğini, -15 bar seviyesinde ise en düĢük çimlenme yüzdesine sahip olduğu gözlenmiĢtir. Yapılan bu çalıĢmada, çimlenme yüzdesi diğer incelenen özelliklere oranla kuraklık stresinden daha az etkilenmiĢ ve bu sebepten kuraklığa toleranslı çeĢitlerin taranmasında çimlenme yüzdesinin iyi bir indeks olmayacağı ortaya konmuĢtur.
Almaghrabi (2012), kuraklık stresinin bazı buğday çeĢitlerinde çimlenme ve fide geliĢim dönemlerindeki parametreler üzerine etkisi ile ilgili çalıĢma yapmıĢtır. Kuraklık toleransı için hızlı bir kriter olarak kullanılabilecek parametrelerin anlaĢılması amacıyla, çimlenme ve fide geliĢim dönemlerinde farklı konsantrasyonlarda (0, 60, 120, 180, 240 ve 300 g/l PEG) polietilen-glikol (PEG) 6000 ile kuraklık stresi uyarılmıĢ, dört yerel (Madini, Kaseemi, Yamanei ve Tabokei), dört yeni geliĢtirilen (Sakha 93, Giza 168, Seds12 ve Masr 1) buğday genotiplerinde kullanıldığı belirtilmiĢtir. Çimlenme para-metrelerinden; son çimlenme yüzdesi, günlük ortalama çimlenme, çimlenme indeksi, ortama çimlenme zamanı ve çimlenme hız katsayısı, fide geliĢim parametrelerinden; kök ve sürgün uzunlukları, sürgün yaĢ ve kuru ağırlıkları, kök yaĢ ve kuru ağırlıkları, fide uzunlukları ve kök sayısı incelenmiĢtir. Farklı ozmotik potansiyelin, kök kuru ağırlığı hariç tüm parametreler üzerinde belirgin bir etkisi olduğu ortaya konmuĢtur. Buna karĢılık, kök sayıları hariç olmak üzere, çimlenme ve fide geliĢim para-metrelerinin, kuraklık stresinde toleranslı ve hassas çeĢitlerin belirlenmesinde uygun olabileceği belirtilmiĢtir.
8
Alaei ve ark. (2012), yaptıkları çalıĢmada kurak koĢullar altında ekmeklik buğday genotiplerinde amino asit içeren iki gübrenin yapraktaki prolin miktarına etkisi-ni araĢtırmıĢlardır. Denemede üç farklı ekmeklik buğday genotipine (Kuhdasht, Cross Sabalan ve Chmran) altı farklı uygulama (normal, aminol forte ve fosnutren, kuraklık (PEG-6000), kuraklık (PEG-6000) + aminol forte, kuraklık + fosnutren) kullanılmıĢtır. AraĢtırma sonuçları, genotipler arasında Chamran’ın en yüksek prolin seviyesine sahip olduğu ve stres koĢulları altında uygulanan fosnutren sıvı gübrelerinin en yüksek prolin yoğunluğuna sahip olduğunu göstermiĢtir. Buna bağlı olarak, amino asit içeren biy-olojik gübrelerin, buğdayda kurak koĢullar altında prolin yoğunluğunu arttırdığını öne sürmüĢlerdir.
Ahmadi ve ark. (2012), yirmi ekmeklik buğday geotipinde, dört farklı PEG 6000 dozu (0, -0.4, -0.8, -1.2 MPa) ile yaptıkları araĢtırmada, tohum çimlenme aĢamasında uygun su potansiyelini ve parametreleri belirlemeyi ve toleranslı genotiplerin taranmasında uygun su potansiyelini ve kullanılacak parametreleri belirlemeyi amaçlamıĢlardır. AraĢtırmada kuraklık seviyelerinin tüm ölçüm karakterlerinde azalma gösterdiği, yapılan varyans analizinin genotip x stres etkileĢiminin önemli farklara dik-kat çektiği belirtilmiĢtir. Çimlenme yüzdesi ve çimlenme gücü parametrelerinin ku-raklığa toleranslı genotiplerin seçiminde kullanılabileceğini ortaya koymuĢlardır.
Ahmad ve ark. (2015); çimlenme döneminde 50 buğday genotipinde kontrol ve kuraklık koĢulları altında ( % 7.5 PEG-6000) çıkıĢ yüzdesi, çıkıĢ indeksi, çıkıĢ oranı indeksi, çıkıĢ enerjisi ve göreceli hücre yaralanması (RCI) özelliklerini incelenmiĢlerdir. En güçlü korelasyon çıkıĢ indeksi ve yüzdesi arasında gözlemlenirken, çimlenme özel-likleri ile RCI arasında negatif iliĢki olduğunu ortaya koymuĢlardır.
Chachar ve ark. (2016), altı ekmeklik buğday genotipinde (MT-1/13, MT-3/13, Khirman, MT- 4/13, MT-2/13 ve Chakwal-86) farklı su stresi (0, -0.5, -0.75, -1.0 MPa PEG-6000) uygulamarıyla çimlenme ve erken dönemdeki tepkilerini incelemeyi amaçlamıĢlardır. Yapılan çalıĢmadai ozmotik stresin altı buğday genotipinde de tohum çimlenmesini, sürgün ve kök uzunluklarını, yaĢ ve kuru ağırlıklarını azalttığını gözlemlemiĢlerdir.
9
Öztürk ve Korkut (2018), 2 yıl boyunca yürüttükleri çalıĢmada, 15 ekmeklik buğday genotipi üzerinde, kuraklığın kök ağırlığına etkisini ve kökün bazı fizyolojik parametrelere etkisini incelemiĢlerdir. Denemede ana parsellerde sapa kalkma evresin-den fizyolojik olum evresine kadar farklı seviyelerde kuraklık uygulanmıĢtır.
ÇalıĢmada; bitki örtüsü sıcaklığı, klorofil kapsamı, tane dolum süresi, baĢaklanma ve olgunlaĢma kök sayıları, kök ağırlığı, stoma sayısı, stoma hacmi, mumsuluk oranı ile kök ağırlığının bu parametreler ile iliĢkisi incelenmiĢtir. Sonuçlara göre; kuraklık stresinin kök ağırlığını azalttığı en fazla kök ağırlığı kuraklık uygulanmayan koĢullarda ölçüldüğünü bildirmiĢlerdir. Kanopi sıcaklığı kurak koĢullarda en yüksek seviyede iken, kuraklık uygulanmayan durumlarda en düĢük seviyede ölçülmüĢtür. Kök ağırlığı art-tıkça kanopi sıcaklığının düĢük seviyede olduğu gözlenmiĢtir. Kök ağırlığı artart-tıkça özel-likle baĢaklanma öncesi ve tane dolum dönemlerinde yaprakta klorofil miktarının art-tığını belirtmiĢlerdir. AraĢtırma, kök miktarının kurağa dayanıklılıkta önemli bir unsur olduğunu, stoma yapısı, klorofil kapsamı ve kanopi sıcaklığı gibi birçok fizyolojik özel-likleri de etkilediğini ortaya koymuĢtur.
Aslan ve ark. (2018), bu çalıĢmada on iki ekmeklik (Gerek-79, , Pehlivan, Flamura-85, Ġkizce-96, Momtchil, Kıraç-66, Demir-2000, Kenanbey, Bayraktar-2000, Pandas, Tosunbey ve Gün-91) ve on siyez buğdayının (Popülasyon-1, Popülasyon-2, Popülasyon-4, Popülasyon-5, Popülasyon-6, Popülasyon-9, Popülasyon-10, Popüla-syon-11, Popülasyon-14, ve Popülasyon-15), yedi farklı doz kuraklık stresi (0: kontrol, 0.09 M, 9.14 ml: 13.71 ml: 0.17 M, 18.28 ml: 0.25 M, 22.85 ml: 0.34 M, 25.15 ml: 0.43M, ve 27.45 ml: 0.51M) uygulayarak, çimlenme hızı, çimlenme gücü, çimlenme oranı, koleoptil uzunluğu, sürgün ve kök uzunluğu, kök yaĢ ve kuru ağırlığı, sürgün/kök uzunluk oranı özellikleri üzerindeki değiĢimleri incelemeyi amaçlamıĢlardır. Yapılan çalıĢmada, kuraklık stresine kök uzunluğu ve sürgün/kök uzunluğu oranının en duyarlı özellikler olduğunu belirtmiĢlerdir. Siyez popülasyonlarının ekmeklik buğday geno-tiplerine oranla kurak koĢullarda daha az geliĢim gösterdikleri sonucunu ortaya ko-ymuĢlardır.
10
2.2. Tuz Stresi
Öncel ve KeleĢ (2002); 3 ekmeklik buğday (Bezostanya-1, Kıraç-66, Seri-82) ve 3 makarnalık buğday genotiplerinin (Kızıltan-91, Kunduru 414-44 ve Ç.1252) tuz stresine (200 mM) karĢı verdikleri tepkileri incelemeyi amaçladıkları bu çalıĢmada, fide döneminde bulunan genotipleri kontrol grubu ve uygulama grubu olarak ayırmıĢlardır. 5 gün boyunca 200 mM NaCl uygulamasının fidelerin kök ve sürgün büyümesini önemli ölçüde engellendiği, tuz uygulaması ile tüm genotiplerde prolin miktarının arttığını gözlemlemiĢlerdir. ÇalıĢmada klorofil-a,b ve toplam klorofil miktarı tüm buğday geno-tiplerinde önemli derecede azalırken; Seri-82 ve Ç-1252 genotipleri klorofil-a kaybının düĢük olduğu görülmüĢtür. Klorofil a/b oranının tuz dayanıklılığının belirlenmesinde önemli bir parametre olabileceği ortaya koymuĢlardır. AraĢtırmada tüm buğday geno-tiplerinde prolin birikmesi gözlenmekle beraber en fazla prolin değerinin kurağa da-yanıklı Kıraç-66 genotipinde gözlendiğini belirtmiĢlerdir. Ekmeklik buğday genotiple-rinin, makarnalık buğday genotiplerine oranla tuz stresine karĢı daha dayanıklı olduklarını, bu durumun prolin biriktirme yeteneğinin zayıflığı ile ilgili olabileceğini öne sürmüĢlerdir.
Akbari ve ark. (2007), Mahdavi, Pishtaz ve Shiraz dahil olduğu üç ekmeklik buğday genotipinin farklı tuz (0, -0.6 ve -1.2 MPa NaCl) ve oksin (0, 1 ve 2 mg L-1
IAA) konsantrasyonları uygulayarak; çimlenme yüzdesi, radikula ve hipokotil uzun-luğunu, fide yaĢ ağırlıklarını, radikula yaĢ ve kuru ağırlıklarını, fide kuru ağırlıklarınını, hipokotil yaĢ ve kuru ağırlıkları üzerine etkilerini incelemiĢlerdir. Artan NaCl kon-santrasyonları, çimlenme yüzdesini, radikula uzunluğunu, hipokotil uzunluğunu, fide yaĢ ve kuru ağırlıklarını azalttığını fakat hipokotil kuru ağırlığının sadece -0.6 MPa NaCl çözeltisinde arttığını bildirmiĢlerdir. Oksin, hipokotil kuru ağırlığını, sürgün yaĢ ve kuru ağırlığını ve hipokotil uzunluğunu arttırmıĢtır, fakat çimlenme yüzdesi ve radikula uzunluğunu etkilememiĢtir. Pishtaz çeĢidinde diğer çeĢitlere kıyasla, yüksek çimlenme yüzdesi, yüksek radikula uzunluğu ve yüksek hipokotil uzunluğu, Shiraz çeĢidinde yüksek kuru kökçük ağırlığı ve Mahdavi çeĢidinde yüksek sürgün yaĢ ve kuru ağırlığı gözlemlemiĢlerdir. Tuz stresinde tohum çimlenme ve erken döneminin en has-sas dönem olduğunu belirtikleri çalıĢmada, ortalama doz (1 mg L-1
) oksin uygula-masının tuz stresinin negatif etkilerini azaltabileceğini bildirmiĢlerdir.
11
Turan ve ark. (2007) yaptıkları araĢtırmada Çakmak-79 ekmeklik buğday geno-tipinde, üç farklı NaCl (0, 25 ve 50 mM ) doz uygulamasının, prolin, klorofil ve Na, Cl, K gibi iyon içeriklerine etkisini incelemiĢlerdir. ÇalıĢmada, NaCl çözeltisi arttıkça buğday genotipinde, K+
/Na+ oranının ve klorofil miktarının azaldığı; fakat prolin miktarının stres koĢulları altında arttığını gözlemlemiĢlerdir.
Duran ve ark. (2010), makarnalık buğday çeĢitlerinde (Altın, Kunduru-1149, Mirzabey, DH-6 ve DH-8) tuz stresi uygulamalarının (100 ve 200 mM NaCl) ) kök uzunluğu, kuru ağırlık, klorofil a, b ve a/b, toplam klorofil miktarları ile karotenoid miktarı üzerine etkilerini incelemiĢlerdir. Tuz stresi uygulamasıyla kök uzunluğu, kloro-fil a, b, kuru madde ve karotenonid miktarlarının azaldığını gözlemlemiĢlerdir. Buğday fideleri üzerinde tuz stresinin olumsuz etkilerini inceleyen Duran ve ark. (2010), 200 mM NaCl uygulamasında bu olumsuz etkinin daha fazla olduğunu belirtmiĢlerdir. AraĢtırmada DH-6 ve DH-8 genotiplerinin diğer genotiplere oranla tuz stresine daha dayanıklı olduklarını ortaya koymuĢlardır.
Deivanai ve ark. (2011), çimlenme ve erken dönemde Malezya’ya ait iki çeltik kültürünün tuz stresi koĢulları altında prolin etkisi üzerinde çalıĢmıĢlardır. ÇalıĢmada, tuz konsantrasyonlarına (0, 100, 200, 300 ve 400 mM) maruz kalan çeltik tohumlarının çimlenme, kök ve sürgün uzunlukları, klorofil içeriği ve protein içeriği önemli bir Ģekil-de etkilendiğini tespit etmiĢlerdir. Çimlenme oranında yavaĢlama, kök ve sürgün uzun-luklarında, klorofil ve protein içeriklerinde açıkça bir azalma olduğunu ortaya ko-ymuĢlardır. Fakat birtakım çalıĢmalar dıĢarıdan uygulanan prolinin, hücresel osmotik dengeyi düzenlemesi ile tuz stresinin negatif etkisini iyileĢtirdiği ortaya koymuĢtur. Bu çalıĢma, prolin dozları (1 mM, 5 mM ve 10 mM) ile ön iĢlem görmüĢ çeltik to-humlarının ve farklı tuz konsantrasyonlarında yetiĢtirilen çeltik toto-humlarının tuzun ters etkisine karĢı olduğunu ispatlamıĢlardır. 1 mM'lık prolin konsantrasyonu ile iĢlem gören çeltik tohumlarının hücresel aktivitelerin uyarıldığı, 10 mM prolin uygulamasının ise yüksek seviyede tuz stresi (300 ve 400 mM NaCl) altında bitki geliĢimini iyileĢtirmede etkisiz olduğunu gözlemlemiĢlerdir.
12
Vardar ve ark. (2014), beĢ ekmeklik (Pehlivan, Golia, Gönen, Köksal-2000, Sag-ittario) ile 3 makarnalık (Gediz-75,Pınar-2001, AltıntaĢ-95) buğday genotiplerinde, beĢ farklı tuz stresi (kontrol dozu, 3.5, 7.0, 10.5, 14.0 ve 17.5 dS m-1) uygulayarak,
çimlen-me dönemindeki etkilerini incelemiĢlerdir. Tuz konsantrasyonlarının artıĢına bağlı olarak incelenen parametrelerde (kök uzunluk ve kuru ağırlıkları, sürgün uzunluk ve kuru ağırlıkları, su alımı, tuz tolerans indeksi, çimlenme oranı ve çimlenme yüzdesi) önemli derecede azalma gözlemlediklerini kaydetmiĢlerdir. Ekmeklik buğday geno-tiplerinden Köksal-200 ve Sagittario, makarnalık buğday genogeno-tiplerinden Gediz-75 ve Pınar-2001’in diğer genotiplere oranla tuz stresine daha dayanıklı olduklarını bild-irmiĢlerdir.
Kahrizi ve ark. (2014), 10 makarnalık buğday çeĢidinde 3 farklı tuz konsantra-syonu (0, 60 ve 120 mM) uygulayarak tuzluluk stresinin bazı fizyolojik ve fenolojik özellikleri (kök uzama süresi, baĢaklanma zamanı, bayrak yaprağın klorofil içeriği, ol-gunlaĢma zamanı) üzerine etkisini araĢtırdıkları çalıĢmada, bu özellikler arasında tuz stresi dozlarının ve çeĢitler üzerindeki etkileĢimlerini baĢaklanma süresi açısından önemli bulmuĢlardır. 61130 çeĢidi ile PGS çeĢitlerinde tuzluluğun artıĢına bağlı olarak baĢaklanma süresinde artıĢ gösterdiklerini ve 605 buğday çeĢidinin en düĢük, 61130 çeĢidinin ise en yüksek kök uzama seviyesine sahip olduğunu bildirmiĢlerdir.
Benlioğlu ve Özkan (2015), Ankara Üniversitesi Ziraat Fakültesi Tarla Bitkileri laboratuvarında yaptıkları çalıĢmada, iki sıralı arpa çeĢitlerinde (Aydan hanım, Bülbül-89 ve Tarm-92) altı farklı tuz stresi (kontrol, 3, 6, 9, 12 ve 15 g/l NaCl) uygulamasının etkilerini incelemiĢlerdir. Çimlenme hızı 4. günde çimlenmiĢ olan tohumlar sayılarak alınan gözlemle belirlenmiĢ, 8. günde alınan ikinci sayımla kök ve sürgün uzunlukları, kök ve sürgün yaĢ ve kuru ağırlıkları, çimlenme gücü gibi parametreler belirlemiĢler ve bu parametreleri istatistiki anlamda önemli bulmuĢlardır. Tarm-92 iki sıralı arpa çeĢidinin incelenen tüm parametrelerde tuza toleransının daha yüksek olduğunu bild-irmiĢlerdir.
Mahboob ve ark. (2016), kontrollü çevre koĢullarında tuza toleranslı ESW-9525 ve kısmen toleranslı Khirman buğday genotiplerine yapraktan harici prolin uygulanarak tuzluluk etkisine karĢı iyileĢtirici etkisini araĢtırmıĢlardır.
13
Yedi günlük buğday fideleri bir hafta boyunca çeĢitli tuz stresi uygulamalarına (0, 60 ve 120 mM NaCl) maruz bırakılmıĢ ve bir hafta sonra yapraktan prolin (0, 50 ve 100 mM) uygulamıĢlardır. Yapraktan prolin uygulamasının kök ve sürgün uzun-luklarını, sürgün yaĢ ve kuru ağırlıklarını, fotosentetik pigmentleri, K+
içeriklerini ve K+/Na+ oranını önemli ölçüde geliĢtirdiğini bildirmiĢlerdir. Yapraktan prolin uygula-masına ve tuzluluğa tepki olarak her iki genotipte de endojen prolin seviyesinde, glisin-betain, toplam çözünen Ģeker ve toplam fenolik içeriklerinde önemli ölçüde değiĢim gözlemlemiĢlerdir. Yapraktan uygulanan prolin 50 mM ve 100 mM, fizyolojik ve biyokimyasal özellikleri tetikleyen bitki büyümesi için bir uyaran olarak bulunmuĢ an-cak 100 mM prolin, kök ve sürgün uzunluklarını, fide kuru ve yaĢ ağırlıklarını, klorofil a, b içeriklerini, glisin-betain, prolin, toplam çözünen Ģekerler, toplam fenolik, K+
içeri-klerini ve K+/Na+ oranını her iki genotiptede geliĢtirerek, tuzluluğun toksik etkilerini iyileĢtirmede en etkili doz olmuĢtur. Fakat, ESW-9525 tuz stresine daha dayanıklı olduğundan, Khirman genotipine oranla osmotik dengeleme ve osmoprotektan içeriği daha geliĢmiĢtir. Bu bulguların, yapraktan uygulanan prolinin buğday genotiplerini tuz toleransına teĢvik ettiğini doğruladığını bildirmiĢlerdir.
Zahra ve Roghayeh (2016); yaptıkları çalıĢmada Roshan ve Bahar adlı buğday çeĢitlerinde, farklı tuz konsantrasyonları (0, 50, 150, 250 mM NaCl) altında, fide döne-minde prolin içeriğine etkisini araĢtırmayı amaçlamıĢlardır. Deneme bitki geliĢiminin kontrolünü tam sağlayabilmek için hidroponik kültür ortamında yürütülmüĢtür. Tuzlu-luk stresi oluĢturmak amacıyla 0, 50, 150, 250 mM NaCl dozları uygulanmıĢtır. ÇalıĢmada tuz dozları arttıkça kök, gövde ve yapraklarda prolin miktarının arttığı ve bu artıĢ miktarının Bahar çeĢidinde daha önemli olduğu ortaya konmuĢtur. Pearson korela-syon katsayısına göre kök ve gövde prolin miktarı önemli bulunmazken, yapraktaki pro-lin miktarının önemli bulunduğunu belirtmiĢlerdir.
Bhusan ve ark. (2016), tuzluluğa duyarlı ve orta toleranslı iki çeltik çeĢidinde, yapraklardan prolin uygulaması ile antioksidan savunma sistemleri oluĢturarak bitkilerin tuz stresi karĢısındaki direncinin geliĢtirilmesi için çalıĢmıĢlardır. KardeĢlen-me döneminde farklı tuz konsantrasyonları (25, 50 ve 100 mM) ile tuz stresi oluĢturmuĢ ve yapraktan farklı dozlarda prolin (0, 25 and 50 mM) uygulamıĢlardır.
14
Tuz stresinin her iki çeltik çeĢidinin büyümesinde ve veriminde belirgin bir düĢüĢe neden olduğu, her iki çeĢidin 50 ve 100 mM NaCl streslerinde tahıl üretmediği ve prolin uygulamasının bu yüksek tuz streslerinin olumsuz etkilerini telafi edemediğini belirtmiĢlerdir. DıĢarıdan prolin uygulamasının; 25 mM NaCl stresinde her iki çeĢidin de bitki büyümesini devam ettirdiğini, tuza duyarlı çeĢidin tane verimini arttırdığını bildirmiĢlerdir. Prolin uygulamasının tuza toleranslı çeltik çeĢidinde hiçbir özelliğe etki etmemesine rağmen, 25 mM NaCl stresinde tuza duyarlı çeĢidin klorofil içeriğini, hücre içi prolin miktarını ve antioksidan enzimlerin aktivitelerini arttırdığını belirtmiĢlerdir. Yapılan çalıĢmada prolin uygulamasının K+
/Na+ oranlarını koruyarak, antioksidan savunma sistemlerini güçlendirerek, tuza duyarlı çeltik bitkilerinin toleransına izin ver-diğini öne sürmüĢlerdir.
Mahboob ve ark. (2018), buğday genotiplerinin tuzluluğa toleransının hızlı bir Ģekilde tarayabilmeyi amaçladıkları çalıĢmalarında, Bitki Islahı ve Genetik Bölümü, Nükleer Tarım Enstitüsü (NIA), Tando Jam, Pakistan'dan temin edilen 11 buğday çeĢidini kullanmıĢlardır. Tuzluluğa toleranslı kontrol genotipi Kiran-95 dahil olmak üzere her genotipe kontrol, 120 ve 160 mM NaCl dozları uygulamıĢlardır. ÇalıĢmada, bütün büyüme kriterlerinin birbiriyle iliĢkili olduğunu, düĢük Na+
ve geliĢtirilmiĢ K + içeriğinin daha iyi K+
/Na+ oranını ortaya çıkaracağından buğdaydaki tuz toleransının taranması için potansiyel fizyolojik kriter olarak bu oranın kullanılabileceğini be-lirtmiĢlerdir.
Bilgili ve ark. (2018), Karatekin Üniversitesi Orman Fakültesi laboratuvarında yaptıkları çalıĢmada, Karatoprak ve Sagittoria ekmeklik buğday çeĢitleri ile Uluslararası Kurak Alanlar AraĢtırma Merkezi’nden (ICARDA) alınan tuz stresine 1 (toleranslı), 8 (orta duyarlı) ve 20 (duyarlı) ıslah hatları kullanmıĢlardır. Yapılan çalıĢma, belirlenen buğday genotiplerine kontrol, 80 mM ve 160 mM NaCl dozlarını uygulayarak; ortalama çimlenme süresi, çimlenme indeksi, sürgün boyu, sürgün yaĢ ağırlığı, çimlenme güç indeksi, çıkıĢ yüzdesi ve indeksi, fide boyu, kök uzunluğu, fide yaĢ ağırlığı özelliklerine etkilerini incelemiĢlerdir. Artan NaCl dozlarının, ortalama çıkıĢ süresi, çıkıĢ güç in-deksi, çimlenme güç indeksi ve ortalama çimlenme süresi değerlerini arttırarak diğer incelenen özellikleri (sürgün boyu, sürgün yaĢ ağırlığı, fide boyu, kök uzunluğu, fide yaĢ ağırlığı, kökçük uzunluğu) önemli ölçüde azalttığını bildirmiĢlerdir.
15
3. MATERYAL VE METOT 3.1. Materyal
3.1.1. Kimyasal Maddeler
ÇalıĢmada pigment ve prolin içeriğinin belirlenmesinde materyal olarak; Ninhidrin,
Sülfosalisilik Asit, OrtoFosforik Asit % 85, Glasiyel Asetik Asit, L- Prolin,
Toluen,
Aseton kimyasal maddeleri kullanılmıĢtır.
3.1.2. Bitki Materyalleri
ÇalıĢmada yer alan ekmeklik buğday genotiplerine ait bilgiler Çizelge 3.1’de verilmiĢtir.
Çizelge 3.1. Denemede Kullanılan Ekmeklik Buğday Genotipleri
GENOTĠP ÖZELLĠK
Envoy Avustralya
Lucilla ProGen Tohum A.ġ
Lok-1 National Seed Corporation(NSC)
RSP-561 Hindistan
Wafia Caso Tarım
3.2. Metot
3.2.1. Bitki Materyalinin YetiĢtirilme Ortamı
Bu çalıĢma, 2018 yılında Çukurova Üniversitesi Bahçe Bitkileri Bölümü Doku Kültürü Laboratuvarında tesadüf parsellerifaktöriyel deneme desenine göre 3 tekerrürlü olacak Ģekilde yürütülmüĢtür. AraĢtırmada kullanılan tohumlar çimlendirme öncesi % 5’lik sodyum hipoklorit (NaClO) ile 100 ml’lik falcon tüpler içerisinde 10 dakika vorteks cihazında çalkalanarak daha sonra tohumlar 3 defa saf su ile durulanmıĢtır.
16
Çimlendirme ortamı; içerisinde filtre kâğıdı olan 12 cm’lik cam petri kaplarına her genotipten 20’Ģer tohum olacak Ģekilde yerleĢtirilmiĢtir (ġekil 3.1).
ġekil 3.1. Dezenfekte edilmiĢ ekmeklik buğday tohumlarının cam petrilere yerleĢtirilmesi
Deneme 24±1 °C ’lik sabit ortam sıcaklığındaki çimlendirme kabininde 18/6 saat gün uzunluğunda 8 gün süreyle yürütülmüĢtür (ġekil 3.2).
17
3.2.2. Tuz Stresi Uygulamaları
Tuz stresi oluĢturmak amacıyla, her bir uygulama için 250 ml saf su içerisinde, 4 dS/m, 8 dS/m ve 12 dS/m dozlarında NaCl çözeltileri hazırlanıp, petrilere yerleĢtirilen tohumlara 10 ml uygulanarak çimlendirmeye bırakılmıĢtır (Çizelge 3.2).
Çizelge 3.2. Denemede Uygulanan NaCl Çözeltileri
Uygulama NaCl g/L
Kontrol Saf su
4 dS/m 1.6 g
8 dS/m 3.2 g
12 dS/m 7.08 g
3.2.3. Kuraklık Stresi Uygulamaları
Kuraklık stresi uygulaması Michel B.E. ve Kaufmann M.R.(1973) göre, 200 ml saf su içerisinde polietilen glikol (PEG 6000) çözeltileri kullanılarak, -0.2 MPa, -0.4 MPa ve -0,6 MPa dozlarında hazırlanmıĢtır (Çizelge 3.3). Hazırlanan çözeltilerden her bir petriye 10 ml uygulanmıĢtır (ġekil 3.3).
Çizelge 3.3. Denemede Uygulanan PEG Çözeltileri
Uygulama PEG-6000 g/L
Kontrol Saf su
-0.2 MPa 119.57 g -0.4 MPa 178.34 g -0.6 MPa 223.66 g
18
ġekil 3.3. Cam petrilere yerleĢtirilen tohumlara PEG çözeltisi uygulaması
3.2.4. Ġncelenen Özellikler ve Belirlenmesi
1. Çim Kını Uzunluğu (cm): Çimlendirmenin 8. gününde, tohum ile ilk yaprağın çıktığı mesafe kumpas ile ölçülmesiyle belirlenmiĢtir (ġekil 3.4).
2. Sürgün Uzunluğu (cm): Çimlendirmenin 8. gününde, sürgünün tohuma bağlandığı yerden yaprağın en uç noktasına kadar olan mesafe kumpas yardımı ile ölçülerek belirlenmiĢtir.
3. Kök Uzunluğu (cm): Çimlendirmenin 8. gününde, kökün tohum ile bağlandığı yerden en uzun kökün uç noktasına kadar olan mesafe kumpas yardımı ile ölçülerek ile belirlenmiĢtir.
4. Sürgün YaĢ Ağırlığı (g): Çimlendirmenin 8. gününde, sürgünlerin tohuma bağlandığı yerden ayırarak hassas terazide tartılması ile bulunmuĢtur.
5. Kök YaĢ Ağırlığı (g): Çimlendirmenin 8. gününde, kökün tohuma bağlandığı yerden ayırarak hassas terazide tartılması ile bulunmuĢtur.
19
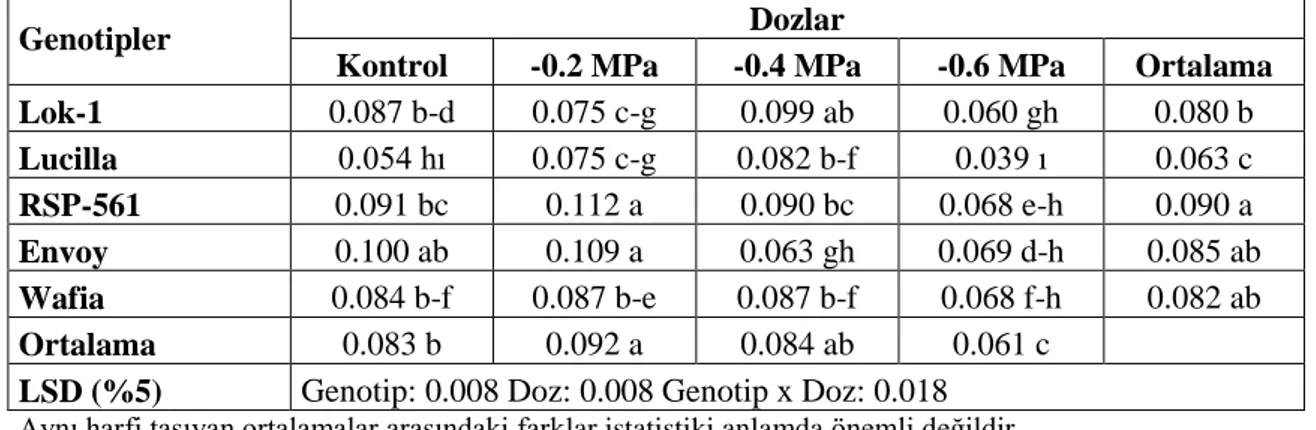
6. Sürgün Kuru Ağırlığı (g): YaĢ ağırlığı ölçülen sürgünler, 70 0C’lik etüvde 48 saat kurutulduktan sonra hassas terazide tartılarak bulunmuĢtur.
7. Kök Kuru Ağırlığı (g): YaĢ ağırlığı ölçülen köklerin, 70 0C‘lik etüvde 48 saat kurutulduktan sonra hassas terazide tartılarak bulunmuĢtur.
8. Çimlenme Hızı (%): Çimlendirmenin 3. günü çimlenen tohum sayısının yüzde değeri olarak hesaplanmıĢtır (ġekil 3.5).
9. Çimlenme Gücü (%): ÇalıĢmanın 8. Gününde, çimlendirmeye bırakılan to-humların, çimlenen tohum sayısının yüzde değeri olarak hesaplanmıĢtır.
10. Turgor Ağırlığı (g): Çimlendirmenin 8. gününde, seçilen sürgünlerden biri 30 ml’lik saf suda 3 saat bekletildikten sonra hassas terazide tartılarak belir-lenmiĢtir.
11. Oransal Su Ġçeriği (RWC): Turgor ağırlığı ölçülen ve kuru ağırlıkları hesaplanan sürgünlerden elde edilen veriler, Smart and Bingham, (1974) be-lirlediği formüle göre hesaplamıĢtır.
RWC = (Sürgün YaĢ Ağırlık – Sürgün Kuru Ağırlık)/(Turgor Ağırlık – Sürgün Kuru Ağırlık)
ġekil 3.4. Denemenin 8. gününde çimlenen buğdayların kumpas yardımı ile ölçümlerinin yapılması
20
ġekil 3.5. Çimlendirme denemesinde her gün gözlem alınması
12. Pigment Ġçeriğinin Belirlenmesi
Pigment içeriği, Arnon (1949)’un belirttiği metoda göre tespit edilmiĢtir. Bu amaçla, 8. gün alınan 0.1 g yaprak örneği, % 80’lik 10 ml aseton içerisinde homojenize edilerek 5 dakika 14 000 rpm’de santrifüj edildikten sonra tüpün üst fazı kullanılmıĢtır (ġekil 3.6). Bitki ekstraktlarının absorbansları, Thermo Scientific Multiskan Go model spektrofotometrede 470, 646 ve 663 nm’de ölçülmüĢtür. Lichtenthaler ve Wellburn (1983) metoduna göre hesaplama yapılmıĢtır. (A: absorbans değeri)
Klorofil a = 12.21*A663-2.81*A646 Klorofil b = 20. 13*A646-5.03*A663
21
ġekil 3.6. Pigment içeriğinin belirlemek için bitki materyali ekstraksiyon iĢlemi
13. Prolin Ġçeriğinin Belirlenmesi
Bates ve ark. (1973)’ın metodu kullanılarak prolin içeriği belirlenmiĢtir. Bunun için 8. gün alınan 0.25 gram yaprak örnekleri, 100 µl % 3 sülfosalisilik asit çözeltisiyle ekstrakte edilmiĢtir. Bitki ekstraktları 5 dakika 14 000 rpm’de santrifüj edildikten sonra, 100 µl üst fazdan alınmıĢ ve üzerine 200 µl asit ninhidrin çözeltisi (0.625 g ninhidrin, 15 ml glasiyel asetik asit ve 10 ml 6M ortofosforik asit ile hazırlanan çözelti, 24 saat +4 °C’de karanlıkta saklanmıĢtır), 200 µl glasiyel asetik asit ve 100 µl sülfosalisilik asit eklenerek 96°C’lik su banyosunda 1 saat inkübe edilmiĢ ve daha sonra örnekler buz banyosuna konularak oda sıcaklığına getirilmiĢtir. Bu aĢamalardan sonra, karıĢıma 1 ml toluen eklenmiĢ ve karıĢımın pembe üst fazından 250 µl alınarak, örneklerin absorbansı Thermo Scientific Multiskan Go model spektrofotometrede toluene karĢı 520 nm’de okunmuĢtur. Standart olarak 0, 0.2, 0.4, 0.8 ve 1 mM konsantrasyonlarında prolin standartları hazırlanıp örneklere uygulanan iĢlemlerin aynısı uygulanmıĢ ve prolin içeriği µmol/g cinsinden hesaplanmıĢtır (ġekil 3.7).
22
ġekil 3.7. Prolin içeriğinin belirlenmesinde yapılan iĢlemler
3.2.5. Verilerin Değerlendirilmesi
AraĢtırmadan elde edilen verilerin varyans analizi, tesadüf parselleri faktöriyel deneme desenine göre JMP 13.0 paket programı kullanılarak % 5 LSD göre yapılmıĢtır.