Partial Characterization of CMCase, Xylanase and Lichenase Enzymes Produced
by Newly Isolated Bacillus sp. CXL
Makbule BAYLAN
1 *Bahri Devrim OZCAN
2Numan OZCAN
31
Çukurova University, Faculty of Fisheries, Department of Basic Sciences, Adana, TURKEY
2Osmaniye Korkut Ata University, Faculty of Arts and Sciences, Department of Biology,
Osmaniye, TURKEY
3
Çukurova University, Faculty of Agriculture, Department of Animal Sciences,
Adana, TURKEY
*Corresponding author: E-mail: [email protected]
Abstract: In the study, cellulase, xylanase and lichenase producing bacterium Bacillus sp. CXL was isolated from the
soil samples collected from agricultural greenhouse. The optimum enzyme activities were observed at 50 °C for CMCase and lichenase, whereas at 40 °C for xylanase. On the other hand, optimum pH values for CMCase, xylanase and lichenase were 8.0, 6.0 and 7.0, respectively. CMCase, xylanase and lichenase were showed 71, 63, and 78% residual activity after pre-incubation at 80 °C for 15 min, respectively. The relative residual activities between 40-80 °C were occurred as 84.4, 86.6, and 92.6%, in the same order. Maximum CMCase, xylanase and lichenase productions of isolate were observed after 36, 24, and 12 hours later from inoculation time. All three enzyme activities were stimulated by CaCl2,MnCl2, CoCl2, and MnCl2, whereas inhibited by HgCl2, and FeCl3. While ZnCl2 and
EDTA stimulated the CMCase and xylanase activities, CuCl2 stimulated CMCase and lichenase. If xylanase activity is
accepted as 100%, the activities of CMCase and lichenase in comparison to the xylanase remain 34 and 56%, respectively.
Keywords: Bacillus sp., CMCase, xylanase, lichenase, characterization
Introduction
Cellulose (insoluble fibres of β-1,4-glucan) and hemicellulose (non-cellulosic polysaccharides such as xylans, mannans, and glucans) are major components of plant cell walls (Wolf et al. 1995). Cellulose the most abundant organic molecule of plant biomass and it is also produced by some animals (e.g., tunicates) and a few bacteria (Lynd et al. 2002). β-1,4-Xylans are mainly found in secondary cell walls of plants, the major component of woody tissue (Thomson, 1993). The β-1,3-1,4-glucans (lichenan) are non-starch linear polysaccharides of cell walls in the endosperm of grains such as barley, oats, rye, rice, wheat, and sorghum (Buliga et al. 1986).
Cellulolytic enzymes are produced by many microorganisms which include fungi and a wide range of bacteria (Wood, 1985). A variety of Bacillus species produce cellulases, including strains of B. cereus (Thayer and David, 1978), B. subtilis (Robson and Chambliss, 1984), B. licheniformis (Dhillon et al. 1985). A cellulase enzyme system include three classes of enzymes; CMCases (EC 3.2.1.4), exoglucanases (EC 3.2.1.91) and β-glucosidase (EC 3.2.1.21) which acts in synergy to release glucose as the end product (Kiio et al. 2016).
Xylans are hydrolysed by a group of enzymes which comprise of endoxylanase (β-1,4-D-xylan xylanohydrolase, EC 3.2.1.8) and β-xylosidase (EC 3.2.1.37), produced by many strains of bacteria and fungi (Bernier et al. 1983). Xylanases are used in bleaching of paper pulp, increasing the brightness of pulp, clarification of fruit juice and improving the digestibility of animal feed (Kamble and Jadhav, 2012).
β-(1,3-1,4)-glucanase (lichenase, EC 3.2.1.73) can only hydrolyse β-1,4-glycosidic bonds adjacent to β-1,3-glycosidic bonds inmixed glycosidic linkages of glycan and lichenan (Gao, 2016). Lichenases have been isolated and characterized from a number of microorganisms including Bacillus species, Paecilomyces thermophila, Aspergillus japonicas, and Trichoderma koningii (Gao, 2016). β-1,3-1,4-glucanases are used in the processing of animal feed, control of fungal pathogens, and release of intracellular materials from microbial cells (Pitson et al. 1993).
In the present study, we aimed to isolate the Bacillus strain producing CMCase, xylanase and lichenases, partial characterization of the enzymes and investigation of the potential availability of the enzymes for animal nutrition.
Materials and Methods
Bacterial strain, culture conditions and detection of extracellular enzymes
Bacillus sp. CXL was isolated from the soil samples collected from agricultural greenhouse located in Adana, Turkey. To select the spore-forming Gram-positive bacteria, 1 g of soil sample was diluted with 10 mL of sterilized distilled water and then was incubated at 80 °C for 10 min (Hamilton et al. 1999). The isolates were cultivated in Luria Bertani (LB) medium (for per litre, tryptone 10 g, yeast extract 5 g, NaCl 10 g, pH 7.5) for 12 h at 37 °C with shaking at 200 rpm. The isolates were screened for CMCase activity on LB-agar-CMC plates containing (g L-1) CMC 2, agar 15 (pH 7.5), for xylanase and lichenase activities on same medium containing (g L-1) xylan 1 and lichenan 1 instead of CMC at 37 °C. To detect CMCase, xylanase and lichenase activities, isolates were selected by flooding the agar plates with Congo-red solution (0.1%) for 15 min (Teather and Wood, 1982). Clear haloes around the colonies indicating of CMCase, xylanase and lichenase activities were obtained by treatment of stained agar plates with 1 M NaCl.
Enzyme assay
CMCase, xylanase and lichenase activities were assayed by adding 1 mL enzyme to substrate (2% w/v) in 50 mM sodium phosphate buffer, pH 6.5, and incubating at optimum pH values (8.0, 6.0 and 7.0 for CMCase, xylanase, and lichenase, respectively) for 30 min. The reactions were stopped by addition of 3 mL of 3,5-dinitrosalicylic acid reagent and A540 nm was measured in a Pharmacia
spectrophotometer (Miller, 1959). An enzyme unit is defined as the amount of enzyme releasing 1 mmol of glucose from the substrate in 1 min at optimum temperature.
Effect of pH and temperature on activity
Temperature and pH effects on enzyme activities were assayed at various temperatures ranging from 30 to 100 °C and at pH values ranging from 5.0 to 12.0 for 30 min. Following buffers were used in pH reactions: 50 mM Na-acetate (pH 5.0-6.0), 50 mM Na-phosphate (pH 6.0-7.0), and 50 mM Tris (pH 7.0-12.0).
Effect of temperature on stability
For determination of thermal stability, the enzymes were pre-incubated at temperatures between 40-100 °C for 15 min. The retained enzyme activities were determined under standard enzyme assay conditions.
Effect of various chemicals on enzyme activities
The effect of various chemicals on the activity of CMCase, xylanase and lichenase were determined by pre-incubating the extracellular enzymes in the presence of CaCl2, MgCl2, ZnCl2, CoCl2,
FeCl3, CuCl2, MnCl2, HgCl2, and EDTA (1 mM each) for 30 min at optimum temperature values before
adding the substrates. Relative enzyme activities were measured at standard enzyme assay conditions (Faulet et al. 2006; Lo et al. 2001).
Results and Discussion
Gram-positive, spore forming and aerobic bacterium Bacillus sp. CXL was isolated from the soil samples collected from agricultural greenhouse. The bacterium showed CMCase, xylanase and lichenase activities on agar plates including certain substrates.
Effect of pH and temperature on enzyme activities and stabilities
The effect of pH on CMCase, xylanase and lichenase activity was determined (Figure 1A). The analysis of enzymes suggests that the enzymes more active around neutral conditions with optimum pH at 6.0 for xylanase, 7.0 for lichenase and 8.0 for CMCase.
The optimal temperature values of enzymes were found to be 40 °C for xylanase, 50 °C for both CMCase and lichenase at optimal pH values (Figure 1B).
Similar optimal temperature values for CMCase from Paenibacillus terrae ME27-1 (Liang et al. 2014), Bacillus sp. HSH-810 (Kim et al. 2005), Bacillus sp. C-14 (Aygan and Arikan, 2008), Bacillus sp. BP23 (Blanco et al. 1995), and pH values from B. vallismortis RG-07 (Gaur and Tiwari, 2015), B. halodurans C-125 (Akita et al. 2005) and Rhodothermus marinus (Hreggvidsson et al. 1996) were reported. On the other hand, similar optimal temperature values with Bacillus sp. CXL xylanase from Bacillus sp. X13 (Aygan and Arıkan, 2009), B. firmus (Tseng et al. 2002), Bacillus sp. BP7 (Lopez et al. 1998) and optimal pH values from B. subtilis (Sa-Pereira et al. 2002), halophilic eubacterium strain SX15 (Wejse et al. 2003), and B. thermantarcticus (Lama et al. 2004), were reported. For Bacillus sp. CXL lichenase, similar optimal temperature values from Orpinomyces sp. GMLF18 (Çömlekçioğlu et al. 2011), B. circulans WL-12 (Kim et al. 2014), and optimal pH values from Bacillu licheniformis UEB CF (Chaari et al. 2012) and Bacillus licheniformis GZ-2 (Gao, 2016) were reported.
The thermal stabilities of CMCase, xylanase and lichenase enzymes were tested by incubating at various heating periods for 15 min. All three enzymes protected all of their activities after pre-incubation at 40 and 50 °C for 15 min and could retain more than 60% of maximum activity after 15 min of pre-incubation at 80 °C (Figure 2C), which was consistent with previous reports that cellulases from some Bacillus strains were stable at 50 °C (Mawadza et al. 2000; Li et al. 2006; Fan et al. 2007; Lin et al. 2012). These results indicate that our enzymes are slightly thermostable and stability to high temperature, make our enzymes a potential for industrial applications.
Time courses of production of extracellular CMCase, xylanase and lichenase, are shown in Figure 1D. The maximum CMCase, xylanase and lichenase activities were reached in 36, 24, and 12 h of culturing time, respectively.
Effect of various chemicals on enzyme activities
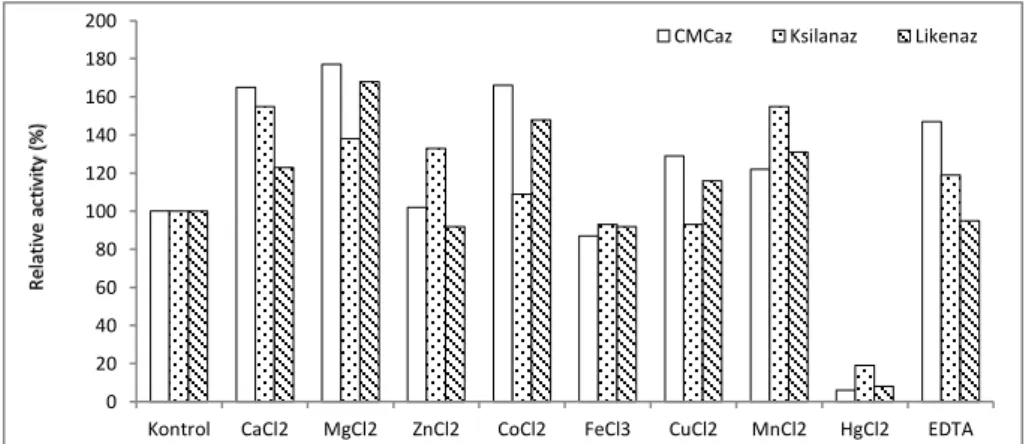
The effect of various chemicals on CMCase, xylanase, and lichenase were investigated by the DNS assay method (Figure 2). Bacterial supernatant was incubated for 30 min with 1 mM of each of the chemicals before determining the residual activity of the three enzymes. Residual activity was determined as the relative amount (%) compared to control (100%).
In this study, activities of the all three enzymes were found to be stimulated by CaCl2 MgCl2,
CoCl2, and MnCl2 with the rates of between 109% and 177%. These results are almost similar to cellulase
from B. subtilis YJ1 (Yin et al. 2010), xylanase from Enterobacter sp. MTCC 5112 (Khandeparkar and Bhosle, 2006), and lichenase from B. circulans WL-12 (Kim et al. 2014). However, in our study, CMCase, xylanase and lichenase activities were strongly inhibited by HgCl2. The strong inhibitory effect of
mercury ion on cellulase (Yin et al. 2010), xylanase (Annamalai et al. 2009), and lichenase (Masilamani, et al. 2013) were also reported previously. An inhibitory effect on enzyme activity by Hg2+ ion usually suggests the reaction with the amino acid cysteine in their side group -SH moiety (Masilamani, et al. 2013).
Figure 1. Effect of varying pH (A), and temperature (B) on the activity of the CMCase, xylanase and lichenase. Effect of varying temperature values on CMCase, xylanase and lichenase stability (C). Effect of
incubation time on enzyme productions by Bacillus sp. CXL (D).
Figure 2. Effect of certain chemical agents on activity of the enzymes
Conclusion
The broad range of temperatures (30 to 70 °C) and pH values (6.0 to 9.0) for high activity and the stability under high temperatures suggest that the Bacillus sp. CXL enzymes may be good candidates for use in industrial applications.
0 20 40 60 80 100 120 5.0 6.0 7.0 8.0 9.0 10.0 11.0 12.0
CMCase Xylanase Lichenase
A R el at iv e a ct iv it y (% ) pH 0 20 40 60 80 100 120 30 40 50 60 70 80 90 100 CMCase Xylanase Lichenase
B R el at iv e a ct iv it y (% ) Temperature (°C) 0 20 40 60 80 100 120 40 50 60 70 80 90 100 CMCase Xylanase Lichenase
Temperature (°C) R el at iv e a ct iv it y (% ) C 0 20 40 60 80 100 120 0 12 24 35 48 60 72 CMCase Xylanase Lichenase
R el at iv e a ct iv it y (% )
Incubation time (hours)
D 0 20 40 60 80 100 120 140 160 180 200
Kontrol CaCl2 MgCl2 ZnCl2 CoCl2 FeCl3 CuCl2 MnCl2 HgCl2 EDTA CMCaz Ksilanaz Likenaz
R el at iv e a ct iv it y (% )
References
Akita, M., K. Kayatama, Y. Hatada, S. Ito, K. Horikoshi, 2005. A novel β-glucanase gene from Bacillus halodurans C-125. FEMS Microbiology Letters. 248:9-15.
Annamalai, N., R. Thavasi, S. Jayalakshmi, T. Balasubramanian, 2009. Thermostable and alkaline tolerant xylanase production by Bacillus subtilis isolated from marine environment. Indian Journal of Biotechnology, 8: 291-297. Aygan, A., B. Arikan, 2008. A new halo-alkaliphilic, thermostable endoglucanase from moderately halophilic Bacillus
sp. C14 isolated from Van soda lake. International Journal of Agricultura and Biology, 10:369-374.
Aygan, A., B. Arıkan, 2009. Production and characterization of multifunctional endoxylanase by Bacillus sp. X13. Turkish Journal of Biology, 33: 231-237.
Bernier, R., D. Michel, L. Jurasek, M.G. Paice, 1983. Isolation and characterization of a xylanase from Bacillus subtilis. Applied and Environmental Microbiology, 46(2): 511-513.
Blanco, A., T. Vidal, J.F. Colom, F.I.J. Pastor, 1995. Purification and properties of xylanase a from alkali-tolerant
Bacillus sp. strain BP-23. Applied and Environmental Microbiology, 61: 4468-4470.
Buliga, G.S., D.A. Brant, G.B. Fincher, 1986. The sequence statistics and solution conformation of a barley (1-3, 1-4)-β-D-glucan. Carbohydrate Research, 157: 139-156.
Chaari, F., F. Bhiri, M. Blibech, S. Maktouf, S. Ellouz-Chaabouni, R. Ellouz-Ghorbel, 2012. Potential application of two thermostable lichenases from a newly isolated Bacillus licheniformis UEB CF: Purification and characterization. Process Biochemistry, 47(3): 509-516.
Çömlekçioğlu, U., E. Özköse, İ. Akyol, F.C. Yazdıç, M.S. Ekinci, 2011., Expression of β-(1,3-1,4)-glucanase gene of
Orpinomyces sp. GMLF18 in Escherichia coli EC1000 and Lactococcus lactis subsp. cremoris MG1363. Turkish
Journal of Biology, 35: 405-414.
Dhillon, N., S. Chhibber, M.A. Saxena, 1985. A constitutive endoglucanase (CMCase) from Bacillus licheniformis-1. Biotechnology Letters, 7(9): 695–697.
Fan, Y.H., W.G. Fang, Y.H. Xiao, X.Y. Yang, Y.J. Zhang, M.J. Bidochka, Y. Pei, 2007. Directed evolution for increased chitinase activity. Applied Microbiology and Biotechnology, 76(1): 135-139.
Faulet, B.M., S. Niamke, J.T. Gonnety, L.P. Kouame, 2006. Purification and biochemical properties of a new thermostable xylanase from symbiotic fungus, Termitomyces sp. African Journal of Biotechnology, 5: 273-282. Gao, Z., 2016. Purification and characterization of a novel lichenase from Bacillus licheniformis GZ-2. Biotechnology
and Applied Biochemistry, 63(2): 249-256.
Gaur, R., S. Tiwari, 2015. Isolation, production, purification and characterization of an organic-solvent-thermostable alkalophilic cellulase from Bacillus vallismortis RG-07. BMC Biotechnology, 15:19, DOI 10.1186/s12896-015-0129-9.
Hamilton, L.M., C.T. Kelly, W.M. Fogarty, 1999. Production and properties of the raw starch-digesting a-amylase of
Bacillus sp. IMD 435. Process Biochemistry, 35: 27–31.
Hreggvidsson, G.O., E. Kaiste, O. Holst, G. Eggertsson, A. Palsdottir, J.K. Kristjansson, 1996. An extremely thermostable cellulase from the thermophilic eubacterium Rhodothermus marinus. Applied and Environmental Microbiology, 62: 3047-3049.
Kamble, R.D., A.R. Jadhav, 2012. Isolation, purification, and characterization of xylanase produced by a new species of Bacillus in solid state fermentation. International Journal of Microbiology, Article ID 683193, 8 pages. Khandeparkar, R., N.B. Bhosle, 2006. Purification and characterization of thermoalkalophilic xylanase isolated from
the Enterobacter sp. MTCC 5112. Research in Microbiology, 157(4): 315-325.
Kiio, I.K., M.F. Jackim, W.B. Munyali, E.K. Muge, 2016. Isolation and characterization of a thermostable cellulase from Bacillus licheniformis strain Vic isolated from geothermal wells in the Kenyan Rift Valley. The Open Biotechnology Journal, 10: 198-207.
Kim, J-Y., S-H. Hur, J-H. Hong, 2005. Purification and characterization of an alkaline cellulase from a newly isolated alkalophilic Bacillus sp. HSH-810. Biotechnology Letters, 27:313-316.
Kim, S-Y., D-B. Oh, O. Kwon, 2014. Characterization of a lichenase isolated from soil metagenome. Journal of Microbiology and Biotechnology, 24(12): 1699-1706.
Lama, L., V. Calandrelli, A. Gambacorta, B. Nicolaus, 2004. Purification and characterization of thermostable xylanase and β-xylosidase by the thermophilic bacterium Bacillus thermantarcticus. Research in Microbiology, 155: 283-289.
Li, Y.H., M. Ding, J. Wang, G.J. Xu, F. Zhao, 2006. A novel thermoacidophilic endoglucanase, Ba-EGA, from a new cellulose-degrading bacterium, Bacillus sp. AC-1. Applied Microbiology and Biotechnology, 70(4): 430-436. Liang, Y-L., Z. Zhang, M. Wu, Y. Wu, J-X. Feng, 2014. Isolation, screening, and identification of cellulolytic bacteria
from natural reserves in the subtropical region of China and optimization of cellulase production by
Paenibacillus terrae ME27-1. BioMed Research International, Article ID 512497, 13 pages.
Lin, L., X. Kan, H. Yan, D. Wang, 2012. Characterization of extracellular cellulose-degrading enzymes from Bacillus
thuringiensis strains. Electronic Journal of Biotechnology, vol. 15, no. 3, DOI: 10.2225/vol15-issue3-fulltext-1.
Lo, H.F., L.L. Lin, H.L. Chen, H.H. Hsu, C.T. Chang, 2001. Enzymatic properties of a SDS-resistant Bacillus sp. TS-23 α-amylase produced by recombinant Escherichia coli. Process Biochemistry, 36: 743-750.
Lopez, C., A. Blanco, F.I.J. Pastor, 1998. Xylanase production by a new alkalitolerant Bacillus. Biotechnology Letters, 20: 243-246.
Lynd, L.R., P.J. Weimer, W.H. Van Zyl, I.S. Pretorius, 2002. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiology and Molecular Biology Reviews, 66(3): 506-577.
Masilamani, R., O.P. Sharma, S.K. Muthuvel, S. Natarajan, 2013. Cloning, expression of -1,3-1,4 glucanase from
Bacillus subtilis SU40 and the effect of calcium ion on the stability of recombinant enzyme: in vitro and in silico
analysis. Bioinformation, 9(19): 958-962.
Mawadza, C., R. Hatti-Kaul, R. Zvauya, B. Mattiasson, 2000. Purification and characterization of cellulases produced by two Bacillus strains. Journal of Biotechnology, 83(3): 177-187.
Miller, G.L., 1959. Use of dinitrosalicylic acid for determination of reducing sugar. Analytical Chemistry, 31: 426-428. Pitson, S.M., R.J. Seviour, B.M. McDougall, 1993. Noncellulolytic fungal β-glucanases: their physiology and
regulation. Enzyme and Microbial Technology, 15: 178-192.
Robson, L.M., G.H. Chambliss, 1984. Characterization of the cellulolytic activity of a Bacillus isolate. Applied and Environmental Microbiology, 47(5): 1039–1046.
Sa-Pereira, P., A. Mesquita, J.C. Duarte, M.R.A. Barros, M. Costaferreira, 2002. Rapid production of thermostable cellulase-free xylanase by a strain Bacillus subtilis and its properties. Enzyme and Microbial Technology, 30: 924-933.
Teather, R.M., P.J. Wood, 1982. Use of Congo-red polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Applied and Environmental Microbiology, 43: 777-780. Thayer, D.W., C.A. David, 1978. Growth of seeded cellulolytic enrichment cultures on mesquite wood. Applied and
Environmental Microbiology. 36(2): 291–296.
Thomson, J.A., 1993. Molecular biology of xylan degradation. FEMS Microbiology Reviews, 104: 65-82.
Tseng, M-J., M-N. Yap, K. Ratanakhanokchai, K.L. Kyu, S-T. Chen, 2002. Purification and characterization of two cellulase free xylanase from an alkaliphilic Bacillus firmus. Enzyme and Microbial Technology, 30: 590-595. Wejse, P.L., K. Ingvorsen, K.K. Mortensen, 2003. Xylanase production by a novel halophilic bacterium increased
20-fold by response surface methodology. Enzyme and Microbial Technology, 32: 721-727.
Wolf, M., A. Geczi, O. Simon, R. Borriss, 1995. Genes encoding xylan and β-glucan hydrolysing enzymes in Bacillus
subtilis: characterization, mapping and construction of strains deficient in lichenase, cellulose and xylanase.
Microbiology, 141: 281-290.
Wood, T.M., 1985. Properties of cellulolytic enzyme systems. Biochemical Society Transactions, 13: 407-410. Yin, L-J., H-H. Lin, Z-R. Xiao, 2010. Purification and characterization of a cellulase from Bacillus subtilis YJ1. Journal of