Başlık: Plasmid-mediated quinolone resistance in Escherichia coli strains isolated from animals in TurkeyYazar(lar):MÜŞTAK, H. Kaan ; İÇA, Tuba ; ÇİFTCİ, Alper ; DİKER, K. SerdarCilt: 59 Sayı: 4 Sayfa: 255-258 DOI: 10.1501/Vetfak_0000002535 Yayın Tarihi:
Tam metin
Şekil

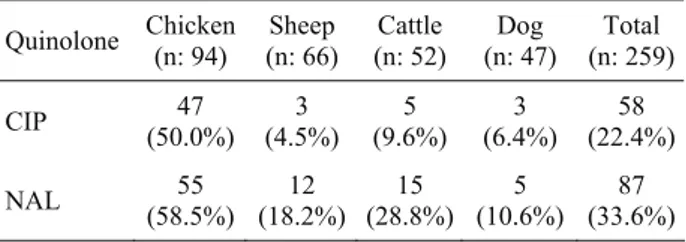
Benzer Belgeler
Based on the literature review, in our study, an intestinal I/R-based stress model was created to evaluate the effect of chronic periph- eral nesfatin-1 application on plasma
[r]
Sağ kulak için elde edilen akustik refleks ölçüm sonuçları incelendiğinde ise, 500 Hz, 1000 Hz ve 2000 Hz frekanslarında saptanan akustik refleks eşikleri açısından
ÖZET: Bu çalışmada Hacettepe Üniversitesi Erişkin Hastanesi’nde hastane enfeksiyonu etkeni olarak izole edilen Escherichia coli, Klebsiella pneumoniae ve
This study uses five antibiotics against Escherichia coli, the results of sensitivity test resistance of Escherichia coli to antibiotics showed that the
Study of Antibiotic Resistance Pattern and Mutation in Genes gyrA and parC of Escherichia Coli Causing Urinary Tract
Similarly in Eksi et al.’s study they found a statistically significant difference of cMLS B resistance in MRSA compared to MSSA but no statistically significant difference
coli isolates included in the study, at least four-fold decrease in MIC values was observed in eight, four and three isolates for ciprofloxacin only, nalidixic acid only and
