Karbonik anhidraz enziminin koyun kalp dokusundan saflaştırılması, karakterizasyonu ve yeni sentezlenen sülfonamid kalkonların etkilerinin araştırılması
Tam metin
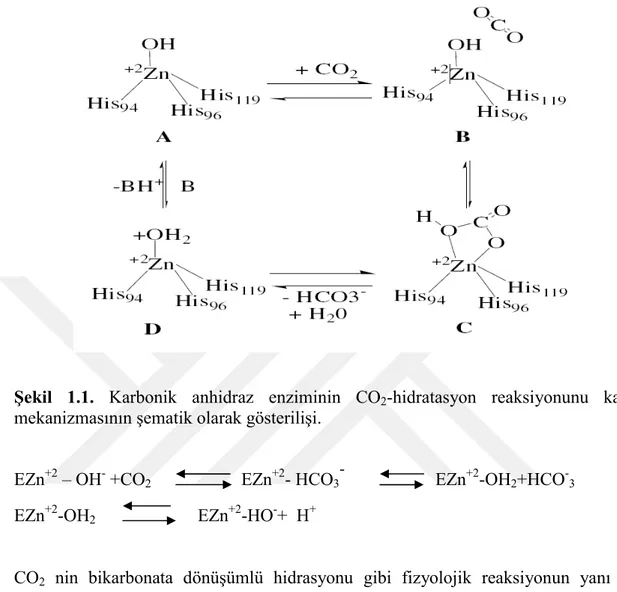
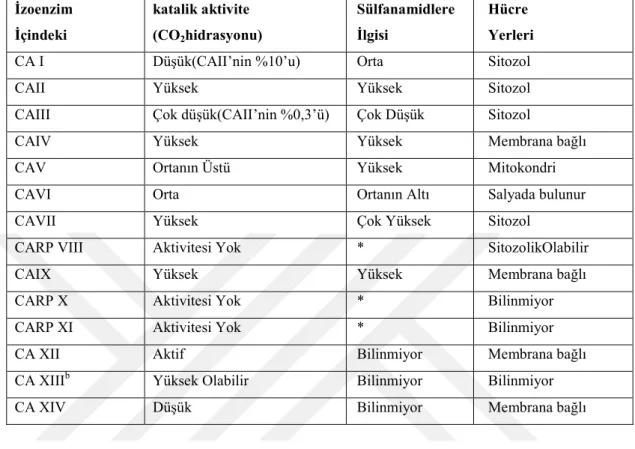
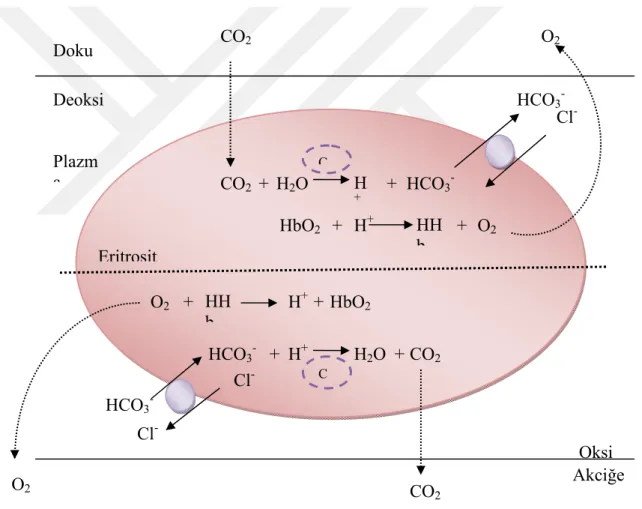
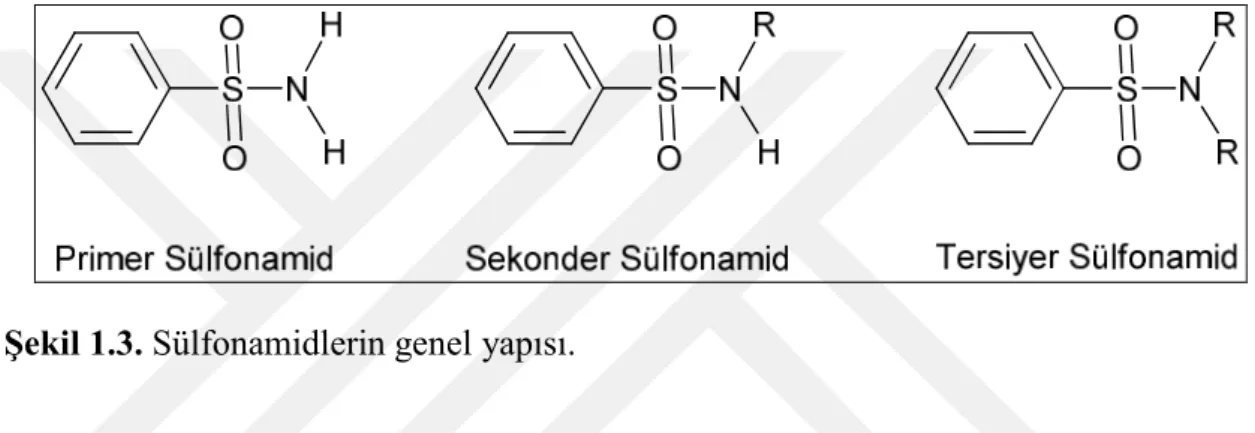
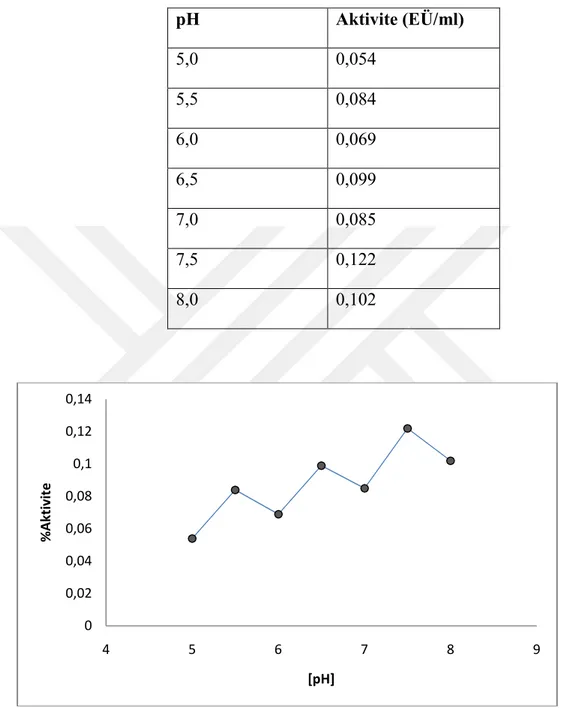
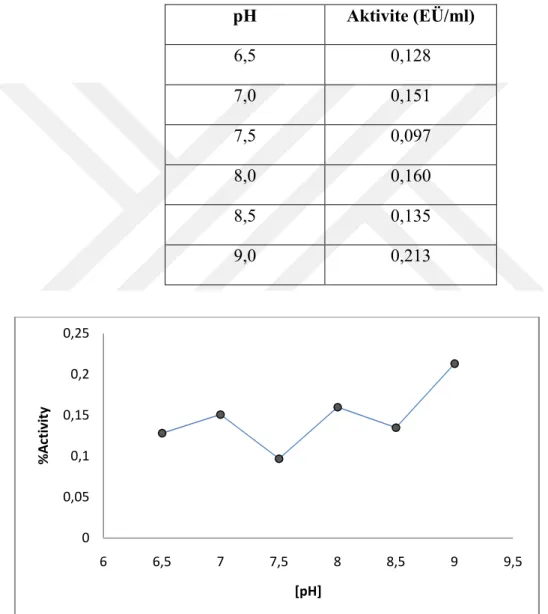
Şekil
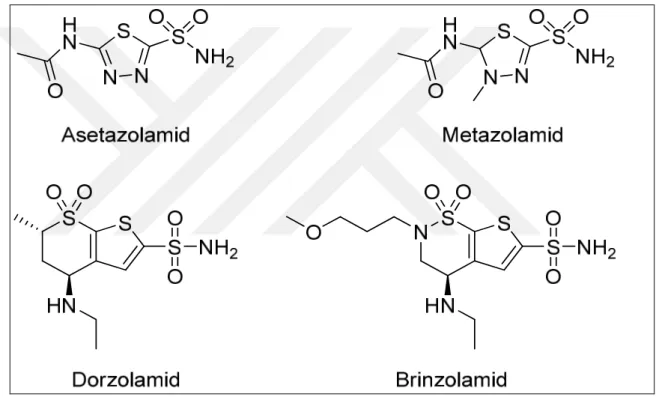
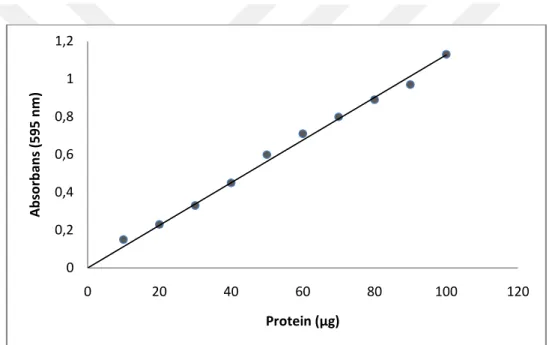
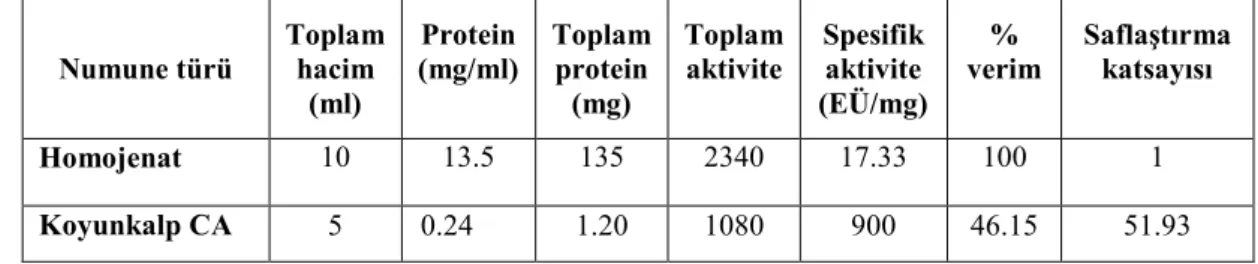
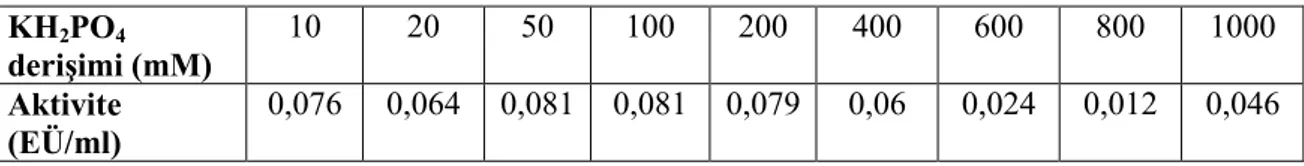
Benzer Belgeler
Plasma disulphide and thiol ratio pairwise comparisons showed that the plasma disulphide/native thiol and disulphide/ total thiol ratios were significantly lower (p<0.001),
Comparison of the specific strength of the as received pure aluminum with the monolithic Al and the Al/ Al 2 O 3 –B 4 C nano-composite shows that, the ARB process increases.. the
Department of Physics and Astronomy, University of British Columbia, Vancouver, BC V6T-1Z1, Canada 2 Department of Physics, Sharif University of Technology, Tehran 11365-9161, Iran
The PMO theory of the secondary and tertiary structure of proteins (i) Electronic configuration of the peptide amide bonds and conformational and H- bonding propensity of
We have also performed quasi-real time SHPM imaging to study how the vortices penetrate into the BSCCO superconductor. We first degaussed the system at 110 K and then applied
sınıf Fen ve Teknoloji dersi programında öngörülen kavramların kazandırılma düzeyini saptamak için, bilgi düzeyi, kavrama düzeyi ve toplam erişi düzeyi
In order to investigate the game effects on attitude change about proximate scenarios (i.e., Israeli–Palestinian scenario for Israeli-Jewish and Palestinian participants and
Simutenkov, bu düzenlemelerin bir tarafta Avrupa Birliği Üye Devleti veya Avrupa Ekonomik Alanı vatandaşları ve diğer tarafta üye olmayan ülkelerin
