Nigella sativanın ratlarda deneysel karbontetraklorür hepatotoksisitesi modelinde koruyucu etkilerinin araştırılması
Tam metin
Şekil
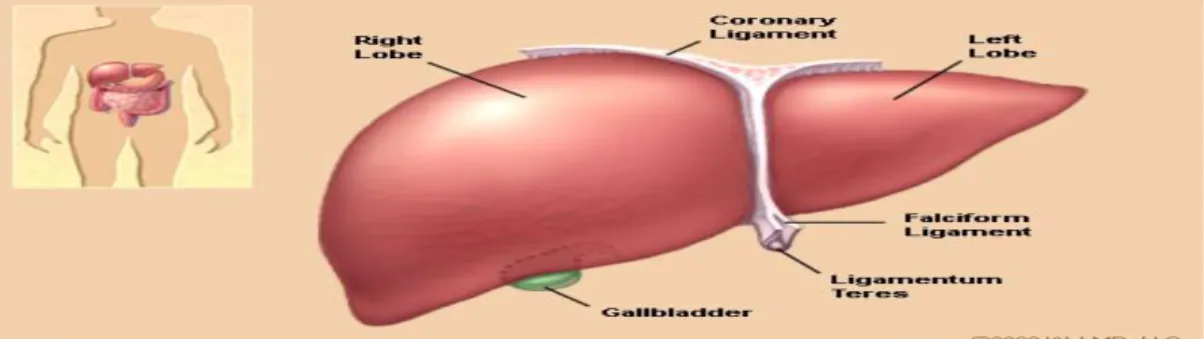
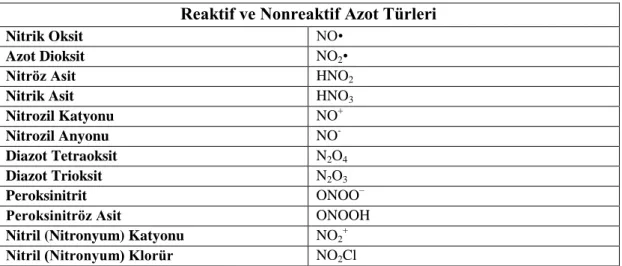
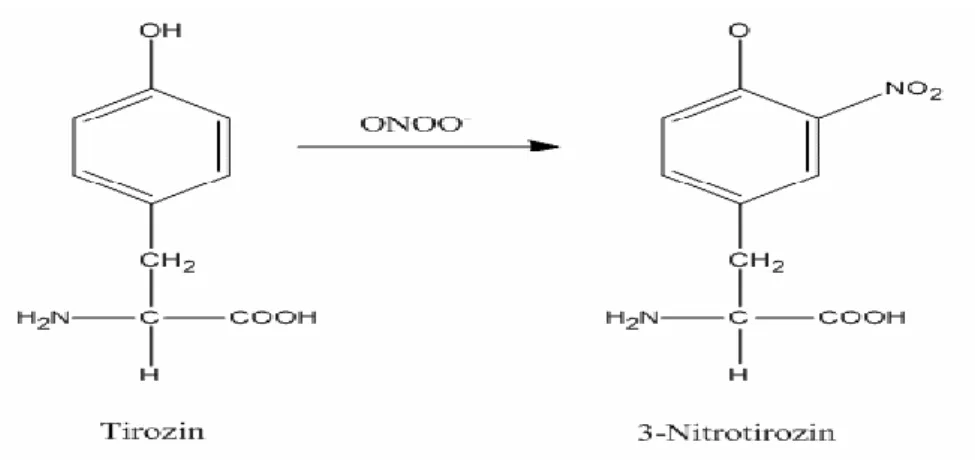
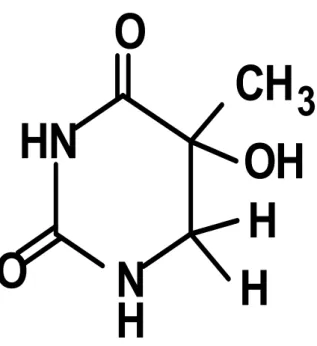
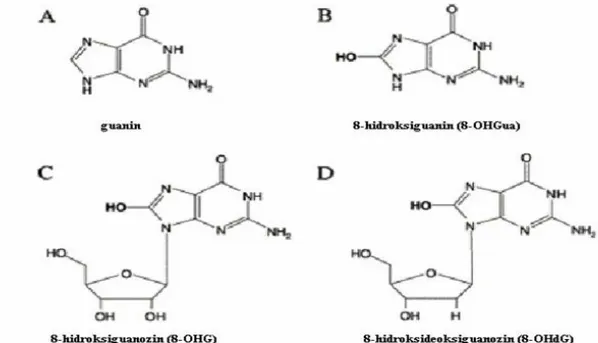
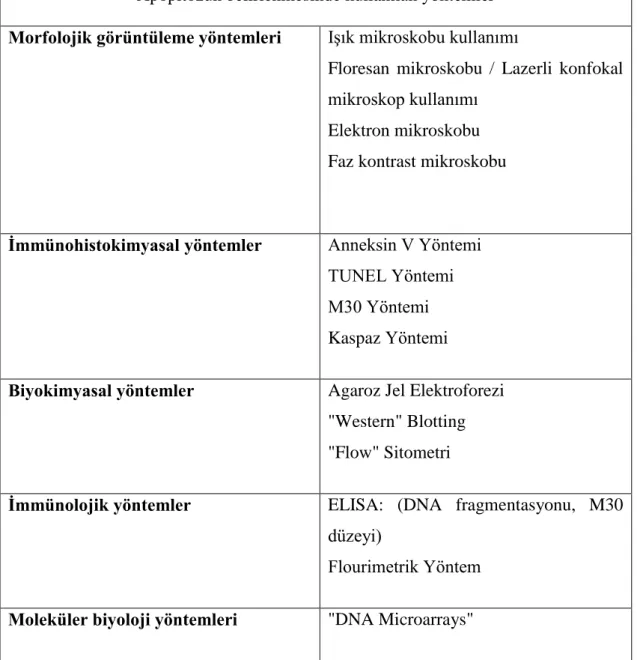
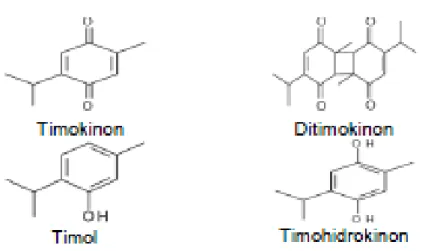
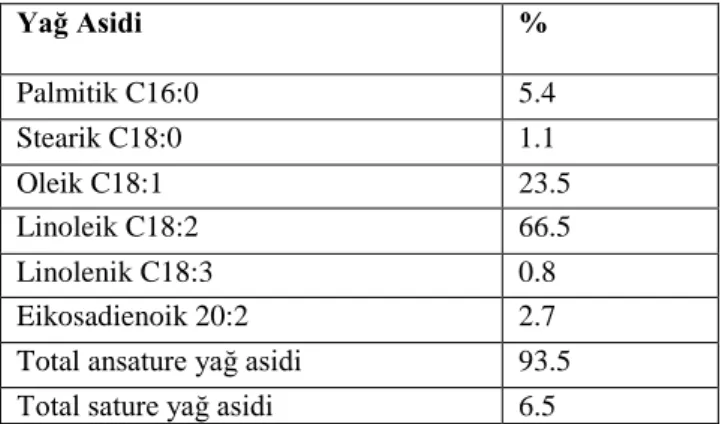

Benzer Belgeler
Hul’de Bedel: Nehaî’ye göre hul’un az ya da çok mal mukabilinde yapılması caizdir. Kocanın hul’ bedeli olarak, verdiği mehirden daha çok veya daha az
Kapitalist zihniyet ve kapitalist üretim anlayışı, orta yaş ve orta yaş üstü, eğitimli, belli bir gelir düzeyine sahip kadın ve erkeklerin yaşlılığı ve
Ankette, sürgündeki ya da merkezi yurtdışında olan haber odaları için (Ahvalnews.com, Ozguruz.org, Artigercek.com) içerik ürettiklerini belirten serbest
Ülkemizde pankreatitin insidansı ile ilgili bir çalışma yapılmamakla birlikte ABD’de yapılan bir çalışmada; ABD’de her yıl AP bağlı olarak yaklaşık 4000
Tropiklerde yetişen, tek cinsi ve 100 kadar türü olan, kışın yaprak dökmeyen ağaç veya çalılardır.. Meyvası drupa tipinde olup tohumu renkli bir arillus ile
of pediatric outcome after prenatal diagnosis and expectant antenatal management of congenital cystic adenomatoid malformation. Hammond PJ, Devdas JM, Ray B, Ward-Platt
In the domestic type hybrid system, the storage of the electricity from the wind turbine and solar panels is achieved by control card, measurement card and connection card is show in
In the present study, 536 rodents were captured from Burdur, Bartin, Giresun, and Yozgat provinces of Turkey between the years 2010 and 2012, and blood samples were examined for
