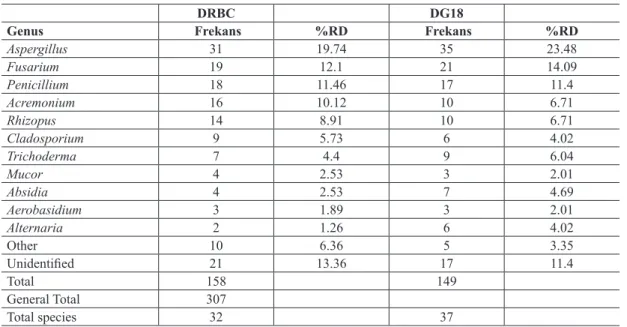
Comparison of two medium according to mould enumeration and recovered species from wheat and feed
Tam metin
Şekil


Benzer Belgeler
History of Conservation biology, Concept of Biodiversity, IUCN threat categories (specis level), Threats over biodiversity, Invasive species and their influence, Conservation at
With regard to the videoing process, Luoma (2004: 39) highlights the advantages of recording the discussion, as they may be used in self reflection of speaking skills. However,
“Kapasiteler ve Kapsayıcılık” başlığı altında ise “kapasite” kavramının, en az demokrasi ve eşitlik kadar kalkınma olgusu için önemli olduğu
Mycotoxin, are toxic substances or metabolites that are synthesized by various pathogenic fungi species and cause intoxications of latent, acute or chronic
Views of Medicine Faculty Academic Staff to Medical Ethics Education (Gazi University Case). Traner CB, Tolchin DW, Tolchin B. Medical Ethics Education for Neu- rology
Şinasî hakkın da monografisi ve Küçük Tarih-i Os- mani, Küçük Taıih-i Islâm, Osmanlı Ta rihi, Resimli ve hantalı Osmanlı Ta rihi, İki Hatırat, Üç
Chow JCY: The Chow technique of endoscopic release of the carpal ligament for carpal tunnel syndrome: 4-years of clinical results.. Chow JCY, Hantes ME, Vernon IL: Endoscopic
nelik çalışmaları ile klasik anaokulu kavra mından temel eğitim dışında tamamen ayrı lan okulda, temel eğitim programını sosyal ve görsel etkinliklerle
