Bitki aktivatörü olarak geliştirilmiş “AkseBio” preparatının toprak kökenli bazı fungal birki patojenlerine etki mekanizmasının araştırılması
100
0
0
Tam metin
(3)
(5)
(7)
(8)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(18)
(19)
(20)
(21)
(23)
(24)
(26)
(27)
(28)
(29)
(30)
(31)
(32)
(33)
(34)
(35)
(36)
Şekil
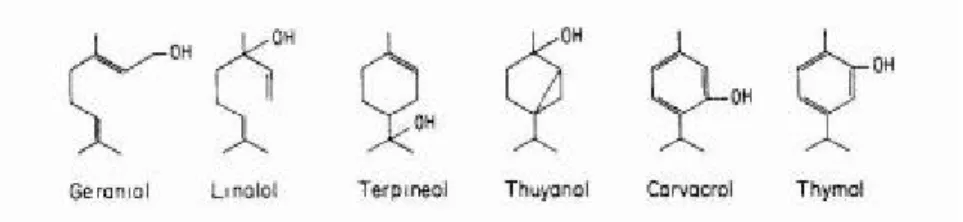
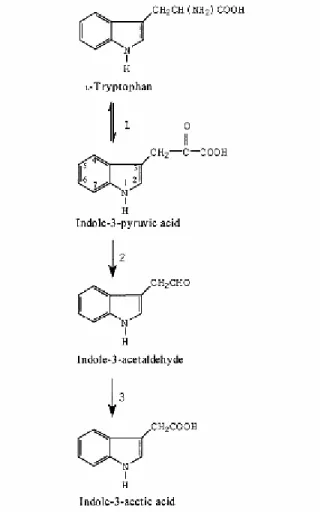
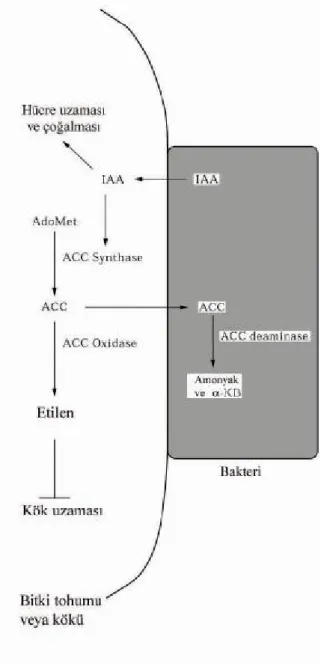
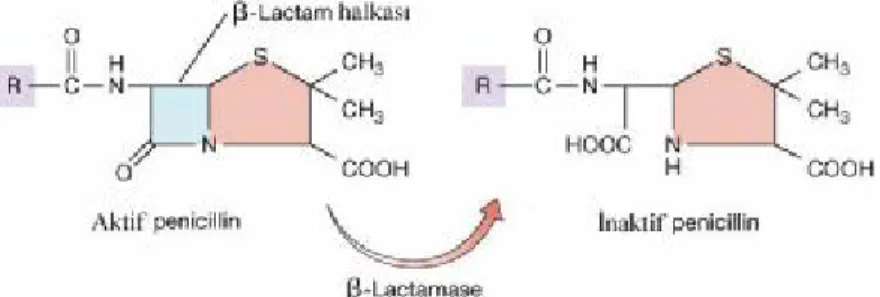
+7
Benzer Belgeler
etmek için kullanılmaktadır, ancak daha uzun zincirli (22-24 karbonlu) yağ asitleri de sentezlenebilmektedir
Yağ asidi açil-CoA, mitokondri iç zarını geçemediği için yapısındaki CoA yerine karnitin bağlanması ile oluşan açil- karnitin, özel bir taşıyıcı
ÇEŞİTLİ KLİNİK ÖRNEKLERDEN İZOLE EDİLEN PSEUDOMONAS AERUGINOSA SUŞLARININ ANTİBİYOTİK DUYARLILIKLARI
Eylül 2012 ile Nisan 2015 tarihleri arasında Necip Fazıl Şehir Hastanesi Mikrobiyoloji Laboratuvarı’na gönderilen çeşitli klinik örneklerden izole edilen toplam
Bu nedenle Ocak 2012-Aralık 2013 tarihleri arasın- da laboratuvarımıza gönderilen çeşitli örneklerden izole edilen Pseudomonas aeruginosa suşlarının antimikrobiyal
Sefalosporinlerin dışında diğer antibiyo- tiklerin de birçoğuna karşı çeşitli mekanizmalar- la yüksek oranda direnç göstermesi ve sahip olduğu direnç profilinin
Çeşitli klinik örneklerden izole edilen Pseudomonas aeruginosa suşlarının isepa- misin ve amikasine duyarlılıkları, Türk Mikrobiyol Cem Derg
Hastane infeksiyonu etkeni Pseudomonas aeruginosa suşlarının antibiyotiklere karşı geliştirdiği çoklu direnç, dünya genelinde önemli bir sorun oluşturmuştur..
Hastanemizde Haziran 2005-Ocak 2010 arasında çeşitli klinik örneklerden izole edilen 570 Pseudomonas aeruginosa izolatının CLSI kriterlerine göre Kirby-Bauer disk difüzyon
