Başlık: Induction of Somatic Embryogenesis from Immature Cotyledons in ‘Tombul’ Hazelnut Yazar(lar):AYGÜN, Ahmet;ŞAN, Bekir;ERDOĞAN, Veli Cilt: 15 Sayı: 2 Sayfa: 113-118 DOI: 10.1501/Tarimbil_0000001080 Yayın Tarihi: 2009 PDF
Tam metin
Şekil
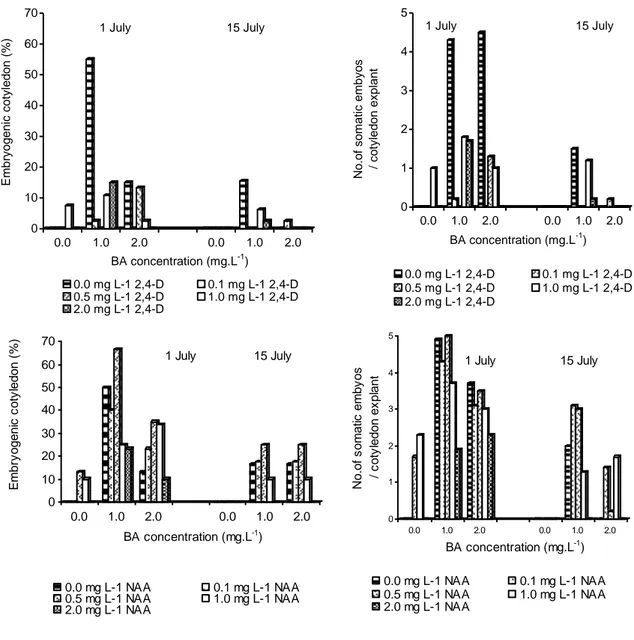
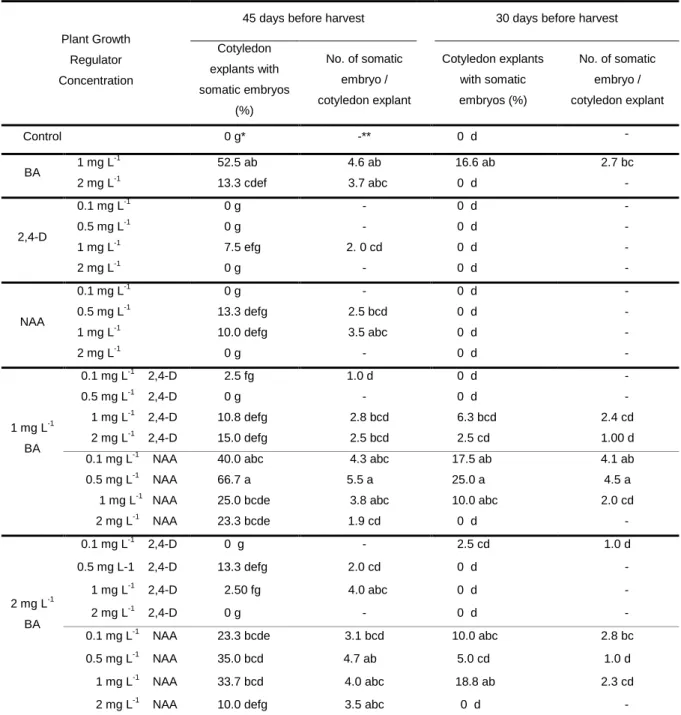
Benzer Belgeler
From a legal point of view, payment systems may be classified into two groups, as either a credit transfer or a debit transfer'' regardless of the means of transmission
Buna göre, Ankara Köy lerinde, köye mahsus konulardan biri olan "boş zamanların değerlen dirilmesi" nden tutunuz da mesken, arazi ve işçilik gücü (labor migra-
ve iğfal ve düşmandan 'ahz-ı sâr ve intikam olunmaksızın ve belki nice kere düşmanı görmeksizin beraberce firar ve külliyen terk-i nâmûs ve 'âr eyledi ğiniz ecilden
olarak çevrilebilecek bu kavramla Sternberger'in kastettiği, kullanımı sakıncalı milliyetçilik kavramının yerine, Alman halkının yarattığı anayasaya bağlı olmak,
Daha önce de değinildiği gibi, ünlü Hun Kağanı Mete Han'dan başlayarak, tüm Türk devletlerinde kağanın kutunu Gök Tanrı'dan aldığına ve O'nun tarafından
Çalışmamızda İM gelişimi için en önemli risk faktörünün 50 yaş üzeri erkek cinsiyet olduğu sonucuna varılmıştır.. Key words: Endoscopy,
He became a fellow of Honeywell Technology Centre in 1982 and a member of the Industrial Advisory Committee of the University of Minnesota in 1986.. Invited by Istanbul
[12] Ahuja, O.P., Planar harmonic univalent and related mappings, Journal of Inequalities in Pure and Applied Mathematics, 6(4) Art. [13] Duren, P., Harmonic Mappings in the
