T.C.
EGE ÜNİVERSİTESİ TIP FAKÜLTESİ
TIBBİ MİKROBİYOLOJİ ANABİLİM DALI
KARBAPENEM DİRENÇLİ KLEBSİELLA PNEUMONİAE SUŞLARINDA KARBAPENEMAZ VARLIĞININ MALDI-TOF MS (MATRIX ASSISTED LASER DESORPTION IONIZATION – TIME OF FLIGHT MASS SPECTROMETRY) İLE ARAŞTIRILMASI VE REAL-TIME PCR YÖNTEMİ İLE KARŞILAŞTIRILMASI
UZMANLIK TEZİ
Dr. Melike Yaşar
DANIŞMAN
Prof. Dr. Fatma Feriha Çilli
İZMİR 2019
T.C.
EGE ÜNİVERSİTESİ TIP FAKÜLTESİ
TIBBİ MİKROBİYOLOJİ ANABİLİM DALI
KARBAPENEM DİRENÇLİ KLEBSİELLA PNEUMONİAE SUŞLARINDA KARBAPENEMAZ VARLIĞININ MALDI-TOF MS (MATRIX ASSISTED LASER DESORPTION IONIZATION – TIME OF FLIGHT MASS SPECTROMETRY) İLE ARAŞTIRILMASI VE REAL-TIME PCR YÖNTEMİ İLE KARŞILAŞTIRILMASI
UZMANLIK TEZİ
Dr. Melike Yaşar
DANIŞMAN
Prof. Dr. Fatma Feriha Çilli
İZMİR 2019
ÖN SÖZ
Tez çalışmamın planlanması, araştırılması, oluşturulması ve yürütülmesinde ilgi ve desteğini esirgemeyen, engin bilgi ve tecrübelerinden yararlandığım sayın hocam Prof. Dr. Feriha ÇİLLİ’ye teşekkürlerimi sunarım.
Teşekkürlerin yetersiz kalacağı hocalarımın da bana dört yıllık üniversite hayatım boyunca kazandırdıkları her şey için ve beni gelecekte söz sahibi yapacak bilgilerle donattıkları için hepsine ayrı ayrı teşekkür ederim.
Ayrıca tezimin istatistiksel analizlerinde yardımcı olan sayın Prof. Dr. Raika DURUSOY’a teşekkür ederim.
Dört yıllık üniversite çalışma yahatımda çalışma ortamımı keyifli hale getiren asistan arkadaşlarım ve çalışma arkadaşlarıma teşekkürlerimi sunarum. Ayrıca tezimin çalışma aşamasında desteklerini hiç esirgemedikleri için Memduh ÖZDEMİR ve Ergül UTKUN’a teşekkür ederim.
Son olarak beni bu günlere sevgi ve saygı kelimelerinin anlamlarını bilecek şekilde yetiştirerek getiren ve benden hiçbir zaman desteğini esirgemeyen bu hayattaki en büyük şansım olan aileme sonsuz teşekkürler.
Melike YAŞAR İZMİR-2019
I İÇİNDEKİLER İÇİNDEKİLER ... I ÖZET ... II ABSTRACT ... IV TABLOLAR LİSTESİ ... VI ŞEKİLLER LİSTESİ ... VII KISALTMALAR LİSTESİ ... VIII
1.GİRİŞ ... 1
2.GENEL BİLGİLER ... 3
2.1. Karbapenemler ... 3
2.2. Gram Negatif Basillerde Antimikrobiyal Direnç Mekanizmaları ... 5
2.2.1. İlacın Hücre İçerisende Etkin Konsantrasyona Ulaşamaması ... 7
2.3.Karbapenem Direnç Gelişimi ... 11
2.3.1.Karbapenemazlar ... 11
2.3.2. Karbapenemaz üretiminin taranacağı fenotipler ... 22
2.4. MALDI-TOF MS ... 24
3.GEREÇ VE YÖNTEM ... 25
3.1. Çalışma Grubu ... 25
3.2. Gradient Test ... 25
3.3. Real Time PCR ... 25
3.4. MALDI-TOF MS Ertapenem Hidroliz Testi ... 26
3.4.1. MALDI-TOF MS analizi ... 27
4.BULGULAR ... 34
4.1. Gradient test sonuçları ... 34
4.2. Real Time PCR sonuçları ... 35
4.3. MALDI-TOF MS ile ertapenem hidroliz testi sonuçları ... 35
5.TARTIŞMA ... 36
6.SONUÇ VE ÖNERİLER ... 40
II ÖZET
Karbapenem dirençli Klebsiella pneumoniae suşlarında karbapenemaz varlığının MALDI-TOF MS (matrix assisted laser desorption ionization – time of flight mass
spectrometry) ile araştırılması ve real-time PCR yöntemi ile karşılaştırılması
Amaç: Karbapenem dirençli Enterobacteriaceae enfeksiyonları tüm dünyada olduğu gibi
ülkemizde de en önemli sağlık sorunlarından birisi olup bu enfeksiyonlarının mortalite ve morbiditesi oldukça yüksektir. Karbapenem dirençli Enterobacteriaceae; DSÖ’nün oluşturduğu, insan sağlığı için ciddi tehdit oluşturan antibiyotiğe dirençli "öncelikli patojenler" listesinin en üst sıralarında yer almaktadır. Bu çalışmada, karbapenemaz aktivitesini kısa sürede saptama amacıyla MALDI-TOF MS’in kullanımına yönelik bir yöntem geliştirilmesi ve geliştirilen yöntemin duyarlılık ve özgüllük değerlerinin belirlenmesi hedeflenmiştir. Ayrıca hastanemizde soyutlanan karbapenem dirençli K. pneumoniae kökenlerinde direnç gen dağılımlarının real time PCR kullanılarak belirlenmesi amaçlanmıştır.
Gereç ve Yöntem: Ocak 2017- Şubat 2019 tarihleri arasında Ege Üniversitesi Hastanesi
Bakteriyoloji Laboratuvar’ında kan kültürlerinden soyutlanıp, stoklanmış olan 74 adet K.
pneumoniae kökeni çalışmaya dahil edildi. Pozitif kontrol olarak Klebsiella pneumoniae
NCTC 13438, negatif kontrol olarak Escherichia coli ATCC 25922 kullanıldı. Kökenlerin öncelikle imipenem, meropenem ve ertapenem gradient test (BioMérieux, Fransa) ile MİK değerleri belirlendi. Daha sonra kökenlerde real time PCR (BD MAX™ CRE Assay, BD Diagnostic Systems, Kanada) ile blaKPC, blaOXA-48 ve/veya blaNDM araştırıldı. Tüm kökenlerde
karbapenemaz aktivitesinin MALDI-TOF MS (BioMérieux, Fransa) ile araştırılmasında ertapenem kullanılarak bir yöntem geliştirildi. PCR ile karşılaştırılarak MALDI-TOF MS ertapenem hidroliz testinin ikinci ve dördüncü saatlerdeki duyarlılık ve özgüllüğü belirlendi.
Bulgular: Gradient test yöntemi ile 74 kökenin 65’i test edilen her üç karbapenem ajana karşı
dirençli, dokuzu ise duyarlı bulundu. Dirençli kökenler için imipenem, meropenem ve ertapenemin hem MİK50 hem de MİK90 değerleri >32 mg/L saptandı. Duyarlı dokuz kökenin
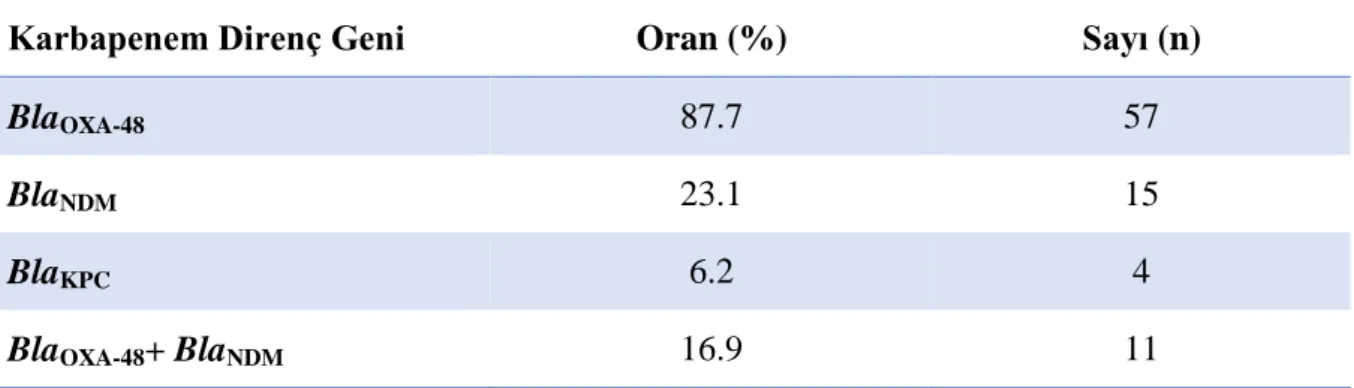
hiçbirisinde blaOXA-48, blaNDM ve blaKPC saptanmadı. Karbapenem dirençli olan 65 kökenin
57’sinde (%87.7) blaOXA-48, 15’inde (%23.1) blaNDM, 4’ünde (%6.2) blaKPC saptandı. 11
III
saatte ertapenemi hidrolize ederken 11 kökende yanlış negatif sonuç elde edildi. Yöntemin ikinci saatteki duyarlılık ve özgüllük oranları sırasıyla %83.1 ve %100 olarak hesaplandı. Dirençli kökenlerin tamamı dördüncü saatte ertapenemi hidrolize etti. Yöntemin dördüncü saatteki duyarlılık ve özgüllük oranı %100 olarak hesaplandı. Karbapenem duyarlı kökenlerin hiçbirisi iki ve dört saatlik inkübasyonlarda ertapenemi hidrolize etmedi.
Sonuç: Çalışmamızda en sık saptanan karbapenemaz tipi %87.7 oranı ile OXA-48 olup bunu
ikinci sıklıkta %23,1 oranı ile NDM izlemektedir. Kökenlerin 11’inde OXA-48 ve NDM birlikte saptandı. Hastanemizde KPC ilk defa bu çalışmada saptandı. Geliştirilen yöntemin ikinci saatteki duyarlılık oranı dördüncü saate göre daha düşük saptandı. İki saatlik inkübasyon sonrası 11 kökende yanlış negatif sonuç elde edildi. Bu kökenlerin tamamında OXA-48 saptandı. Yine bu 11 kökenin üçünde eş zamanlı olarak NDM’nin de bulunduğu belirlendi. Laboratuvarda klinik örneklerden gram negatif bakteriler soyutlandığında; geliştirilen bu yöntem karbapenemaz aktivitesini rutin duyarlılık testlerinden daha kısa sürede aynı gün içerisinde (2-4 saatte) saptamaya olanak sağlamaktadır. Eş zamanlı olarak birden fazla köken çalışılabilir. MALDI-TOF MS ile karbapenemaz aktivitesinin saptanması, henüz gelişme aşamasında olan hızlı, ucuz, umut verici bir yöntemdir.
IV ABSTRACT
Investigation of The Presence of Carbapenemase in Carbapenem-resistant Klebsiella pneumoniae Strains by MALDI-TOF MS (Matrix Assisted Laser Desorption Ionization
– Time Of Flight Mass Spectrometry) and Comparison with Real-Time PCR Method
Aim of the study: The increasing prevalence of bacterial resistance to antibiotics is an
important public health problem. Infections due to carbapenem resistant Enterobacteriaceae strains are among the most important health problems in our country as well as in the world. Infections caused by carbapenem resistant bacteria are associated with significant morbidity and mortality worldwide. Carbapenem-resistant Enterobacteriaceae are among the top tier of the WHO list of antibiotic-resistant “priority pathogens” that pose the greatest threat to human health. The aim of this study is to develop a procedure to detect carbapenemase activity by MALD-TOF MS and it is aim to to calculate the sensitivity and specificity of the developed method. The second aim of this study is to determine the distribution of resistance genes of carbapenem-resistant K. penumoniae strains isolated in our hospital using real time PCR.
Material and methods: Between January 2017- February 2019 K. pneumoniae strains (n=74)
isolated and stocked from blood culture samples in Ege University Hospital Bacteriology Laboratory were included in the study. Klebsiella pneumoniae NCTC 13438 was used as positive control and Escherichia coli ATCC 25922 was used as negative control. First, MIC values of the isolates were determined by imipenem, meropenem and ertapenem gradient test (BioMérieux, France). Then, the strains were investigated by using real time PCR (BD MAX™ CRE Assay, BD Diagnostic Systems, Canada) to detect blaKPC, blaOXA-48 and/or blaNDM carbapenem resistance genes. A method was developed using ertapenem for the
determination of carbapenemase activity by MALDI-TOF MS (BioMérieux, France) of all strains. It was evaluated whether two and four hours incubation of the bacteria in ertapenem solution carried out ertapenem hydrolysis with using MALDI-TOF. Sensitivity and specificity of MALDI-TOF MS were determined by PCR.
Results: 9 strains were susceptible to the imipenem, meropenem and ertapenem. 65 strains
were resistant to imipenem, meropenem and ertapenem. Imipenem, meropenem and ertapenem MIC50 and MIC90 values of all resistant strains were > 32 mg/L. BlaOXA-48, blaNDM
V
resistant to any of the carbapenems, 57 (87.7%) had blaOXA-48, 15 (23.1%) had blaNDM, and 4
(6.2%) had blaKPC. BlaOXA-48 and blaNDM genes were detected together in 11 (16.9%) strains.
The sensitivity and specificity of MALDI-TOF MS at the second hour were 83.1% and 100%, respectively. Sensitivity and specificity at the fourth hour were 100%. MALDI-TOF MS showed that 54 strains resistant to carbapenems hydrolyze ertapenem at second hour; All carbapenem resistant strains were found to hydrolyze ertapenem at the 4th hour. Carbapenem susceptible strains did not hydrolyze ertapenem.
Conclusion: In our hospital, blaOXA-48 and blaNDM were detected 87.7% and 23.1%
respecively. The sensitivity and specificity of MALDI-TOF MS at the second hour were 83.1% and 100%, respectively. Sensitivity and specificity at the fourth hour were 100%. In the MALDI-TOF MS ertapenem hydrolysis test, false negative results were observed at the second hour. These originated from OXA-48-producing origins. Determination of carbapenemase activity by TOF MS is an evolving and promising method. MALDI-TOF MS provides information on the carbapenemase activity of the bacteria in two to four hours. After bacterial identification, it can be used to show carbapenemase activity on the same day. Multiple samples can be tested at the same time. Can be used in the presence of an outbreak. It is an easy test because routine laboratory materials are used in this method. Spectrum analysis is subjective because it is performed manually.
VI TABLOLAR LİSTESİ
Tablo 1. β-Laktamazların Sınıflandırması ... 10
Tablo 2. KPC saptanan türler ... 12
Tablo 3. Ertapenemin doğal ve yıkım ürünlerine ait pik değerleri (m/z) ... 28
Tablo 4. Kökenlerin duyarlılık sonuçları ... 34
Tablo 5. Dirençli kökenlerin duyarlılık sonuçlarına göre MİK50 ve MİK90 değerleri ... 34
VII ŞEKİLLER LİSTESİ
Şekil 1. Karbapenem çekirdek yapıları ve substituentleri ... 4
Şekil 2. MALDI-TOF MS ertapenem hidroliz testi spektrumu ... 29
Şekil 3. MALDI-TOF MS ertapenem hidroliz testi spektrumu. ... 30
Şekil 4. MALDI-TOF MS ertapenem hidroliz testi spektrumu ... 31
VIII KISALTMALAR LİSTESİ
MALDI TOF : Matrix assisted laser desorption ionization time of flight ESI : Elektrospray iyonizasyon
MS : Kütle spektrofotometrisi PCR : Polimeraz zincir reaksiyonu MİK : Minimal inhibitör konsantrasyon CHCA : 4-hydroxy-α-cyanocinnamic asit
CRE : Karbapenem dirençli Enterobacteriaceae YBÜ : Yoğun bakım ünitesi
PBP : Penisilin bağlayan protein
CIM : Karbapenem inaktivasyon yöntemi
ICARE : Yoğun Bakım Antimikrobiyal Direnç Epidemiyolojisi
IMP : İmipenemaz
VIM : Verone integron-encoded metallo-β-laktamaz NDM : New Delhi metallo-β-laktamaz
GIM : German imipenemaz
SPM : Sao Paulo metallo-β-laktamaz
SIM : Seoul imipenemaz
MβL : Metallo-β-laktamaz
PGFE : Pulsed-Field Jel Elektroforez MLST : Multilocus Sequence Typing SPC : Örnek İşlem Kontrolü DHP-I : Dehidropeptidazı EMEA : Avrupa İlaç Ajansı
IBC : Integron-borne sefalosporinaz
ESBL : Genişlemiş spektrumlu betalaktamazlar OMP : Dış membran proteini
IX ABC : ATP-Binding Casette Süper Ailesi
MFS : Major Facilitator Süper Ailesi
SMR : Small Multidrug Resistance Süper Ailesi
MATE : Multidrug and Toxic Compound Extrusion Süper Ailesi RND : Resistance-Nodulation-cell Division
NMC : Not-metalloenzim karbapenemaz SME : Serratia marcescens enzyme
KPC : Klebsiella pneumoniae carbapenemase GES : Guiana extended spectrum
ABD : Amerika Birleşik Devletleri
CDC : Centers for Disease Control and Prevention CLSI : Clinical and Laboratory Standards Institute
1 1. GİRİŞ
Karbapenem dirençli Enterobacteriaceae kökenlerine bağlı enfeksiyonlar tüm dünyada olduğu gibi ülkemizde de en önemli sağlık sorunlarındandır. Hastanelerde genellikle yüksek mortalite ile seyreden nozokomiyal enfeksiyonlar ve salgınlar olarak ortaya çıkmaktadır. Özellikle duyarlılık paterni çoklu ilaca dirençli suşlar olarak karşımıza gelmektedir. Karbapenem dirençli Klebsiella pneumoniae’nın neden olduğu enfeksiyonlar genellikle, yoğun bakım ünitelerinde yatan, altta yatan hastalıkları olan ve uzun süreli antibiyotik tedavisi alma öyküsü olan hastalarda ortaya çıkmaktadır (1). Bu dirençli mikroorganizmalara bağlı enfeksiyonlar hastaların yatış süresini uzatmakta, mortalite, morbidite ve tedavi maliyetlerinde artışa neden olmaktadır (2). Bu enfeksiyonların tedavisinde kullanılabilecek etkin antibiyotik seçeneklerinin sınırlı olması ve direncin hızla yayılabilmesi nedeniyle de önemlidir. Karbapenem dirençli Klebsiella türlerine bağlı sporadik olgular ve salgınlar ülkemizde de artan sıklıkta raporlanmaktadır (3-5).
Gram negatif basillerde karbapenemlere direnç, bir veya birden mekanizma sonucunda ortaya çıkabilir. Bu mekanizmalar karbapenemleri hidrolize eden β-laktamaz üretimi, dış membran porin değişiklikleri ve aktif dışa atım (efflux) pompalarıdır. Hedef penisilin bağlayan protein değişiklikleri genellikle gram pozitif mikroorganizmalarda görülmektedir (6). Karbapenemazlar ile ortaya çıkan direnç, diğer mekanizmalarından daha sık görülmektedir. Bu enzimleri kodlayan genlerin plazmidler üzerinde bulunması, direncin aynı ve farklı türler arasında hızla yayılmasına yol açar. Direncin kolay yayılabilmesi nedeniyle, bu kökenlerin mümkün olan en kısa sürede saptanması, enfeksiyon kontrolu ve tedavinin doğru yönlendirilmesini sağlamakta ve halk sağlığı açısından önem arz etmektedir (2). Albiger ve arkadaşlarının yaptığı bir çalışmada, Türkiye’nin de bulunduğu 38 Avrupa ülkesini kapsayan ve ülkelerin karbapenemaz tip dağılımlarının değerlendirildiği raporda ülkemiz; OXA-48 karbapanemazlar açısından endemik, NDM-1 karbapenemazlar açısından bölgesel yayılım, VIM açısından da hastanelerden salgınlarının bildirildiği bir ülke olarak tanımlanmıştır (7). Karbapenem direnci açısından ülkemiz gibi yüksek orana sahip ülkelerde, bu direncin saptanmasının önemi gittikçe artmaktadır.
Enterobacteriaceae kökenlerinde rutin duyarlılık testlerinde azalmış karbapenem
duyarlılığı saptandıktan sonra, karbapenemaz varlığının araştırılması için fenotipik ve genotipik testler uygulanabilir. Fenotipik yöntemler arasında; kombinasyon disk yöntemi, biyokimyasal (kolorimetrik) testler (CarbaNP testi, Blue-Carba testi, -CARBA testi™), karbapenem inaktivasyon yöntemi (CIM), lateral akım yöntemleri ve yeni bir yöntem olan karbapenem hidrolizinin MALDI-TOF ile saptanması yer almaktadır (8). Moleküler testler
2
duyarlılık ve özgüllükleri yüksek olan hızlı testler olmalarına karşın maliyetlerinin yüksek olması ve teknik altyapı gerektirmeleri nedeniyle fenotipik testlerin kullanımı daha ön plandadır (2).
MALDI-TOF MS farklı kimyasal bileşiklerin molekül ağırlıklarının belirlenmesi için kullanılır. MALDI-TOF MS rutin mikrobiyoloji laboratuvarlarında bakterilerin ve mantarların tanımlanmasında kullanılmaktadır (9). Son dönemlerde MALDI-TOF MS ile hızlı sonuç ve düşük maliyet ile karbapenemaz aktivitesinin saptanması, gittikçe daha da önem kazanmaktadır.
Bu çalışmada, karbapenem dirençli Klebsiella pneumoniae kökenlerinde karbapenemaz aktivitesinin MALD-TOF MS ile araştırılması ve real-time PCR yöntemi ile karşılaştırılması amaçlanmıştır. Bunun yanısıra kökenlerin real-time PCR ile direnç genlerinin araştırılması amaçlanmıştır.
3 2. GENEL BİLGİLER
2.1. Karbapenemler
Karbapenemler, β-laktam antibiyotiklerin en yeni grubunu oluşturan, geniş spektrumlu antibiyotiklerdir. Farklı PBP’lere karşı yüksek afiniteye sahip olmarı, A sınıfı genişletilmiş spektrumlu laktamazlar ve C sınıfı laktamazlar (AmpC'ler) dahil olmak üzere çoğu β-laktamazlara karşı stabil olmaları, gram pozitif ve gram negatif aerobik ve anaerobik bakterilerin dış membranlarından etkin bir şekilde geçebilmeleri sayesinde geniş bir etki spektrumuna sahip ajanlardır (6). Enterobacteriaceae grubunda sefalosporinlere karşı artan direnç nedeniyle dünya çapında imipenem, meropenem, doripenem, ertapenem, panipenem ve biapenem gibi karbapenemler kullanıma girmiştir (10).
Karbapenemler, toprakta bulunan bir mikroorganizma olan Streptomyces cattleya tarafından üretilen bir antibiyotik olan tiyenamisin türevidir. Beş üyeli tiazolidin halkasındaki 1. pozisyondaki kükürt yerine bir karbon atomu ve C2 ile C3 arasında çift bağ içerirler. Bu yapısıyla penisilinlerden farklılık gösterir. Altıncı karbon atomunda trans-1α-hidroksietil yan zinciri sayesinde β-laktamazlara karşı daha stabil bir yapıya sahiptirler (11).
Tiyenamisinin kimyasal olarak kararsız bir yapıya sahip olması, daha stabil yapıya sahip bir molekül arayışına neden olmuştur. İlk olarak N-formimidoil türevi olan imipenem geliştirilmiştir (6, 11). İmipenem ve ardından geliştirilen panipenem daha stabil tiyenamisin türevleridir. Hem İmipenem hem panipenem, memeli böbrek dehidropeptidazı (DHP-I) tarafından in vivo olarak indirgenir ve bu enzimin seçici bir antagonisti olan silastatin veya betamipron ile birlikte kullanılmaktadır (6). İmipenem 1985 yılında, komplike mikrobiyal enfeksiyonların tedavisinde kullanılabilen ilk karbapenem olmuştur. Daha geniş spektrumlu, daha stabil karbapenemlerin keşfedilmesi yolculuğunda şu anda mevcut olan diğer bileşikler; meropenem, biapenem, ertapenem ve doripenemdir (11). Meropenem, ertapenem ve doripenem, ikinci karbon atomunda 1β-metil ve pirolidinil halkasına sahip olarak imipenemden farklıdır. 1β-metil bileşeni, DHP-I enzimine karşı stabilite sağlamaktadır (6). Karbapenemlerin çekirdek yapıları ve substituentleri Şekil 1’de gösterilmiştir.
4 Şekil 1. Karbapenem çekirdek yapıları ve substituentleri (12)
Karbapenemler, yüksek molekül ağırlıklı PBP'lere bağlanarak hücre duvar sentezini inhibe ederler. Bakteri hücre duvarı boyunca kolayca dağılmazlar, periplazmik boşluğa ulaşmak için gram negatif bakterilerin dış zarını, porin olarak da bilinen dış membran proteinleri boyunca geçerler. Periplazmik aralığa ulaşan karbapenemlerin etkisi, bakteri hücre duvarına difüze olarak PBP’lere bağlanması ile başlar (10, 11). PBP'ler, bakteri hücre duvarında peptidoglikanın oluşumunu katalize eden enzimlerdir (transglikozlar, transpeptidazlar ve karboksipeptidazlar). Karbapenemler, PBP'lerin peptidaz bölgesini inhibe ederek etkinlik gösterir. Peptit çapraz bağlanmasını ve ayrıca diğer peptidaz reaksiyonlarını inhibe ederler. Hücre duvar sentezi, yapım ve yıkımın denge halinde olduğu dinamik bir süreçtir. PBP'ler karbapenemler ile inhibe edildiğinde denge, yıkım lehine döner. Sonunda peptidoglikan zayıflar ve hücre ozmotik basınca bağlı olarak patlar (10). Karbapenemlerin, çeşitli bakterilerin farklı PBP'lerine karşı gösterdiği afinite, bu ajanların spektrumunun genişlemesine katkı sağlar (11).
Genel olarak, karbapenemler, enfeksiyöz bakteriler üzerindeki konsantrasyondan bağımsız öldürme etkileri nedeniyle, invaziv veya hayati tehlike taşıyan enfeksiyonların
5
tedavisinde diğer antimikrobiyallere göre tercih edilir. Geniş spektrumludurlar. Anaeroblar da dahil olmak üzere gram pozitif ve gram negatif bakterilere etkilidirler (10).
Karbapenemler arasında meropenem, doripenem, panipenem ve ertapenem gibi pirolidin türevlerine sahip siklik amin tiplerinde daha geniş bir aktivite spektrumu bulunur. Genel olarak, imipenem, panipenem ve doripenem, gram pozitif bakterilere karşı güçlü antibiyotiklerdir. Meropenem, biapenem, ertapenem ve doripenem, gram negatif bakterilere karşı daha etkilidirler. Pseudomonas aeruginosa'ya karşı ertapenemin etkinliği, meropenem veya imipenem ile karşılaştırıldığında nispeten sınırlıdır. Ertapenem, Pseudomonas türlerine ve diğer non-fermentatif gram negatif bakterilere karşı en düşük aktiviteye sahip olan karbapenemdir. Halen, klinik uygulamada antipseudomonal ajanlar olarak doripenem, imipenem ve meropenem gibi farklı tiplerde karbapenemler kullanılmaktadır. Acinetobacter
baumannii imipenem ve doripeneme , meropenemden daha duyarlıdır (11).
İmipenem renal dihidropeptidaz enzimi tarafından yıkılır, bu nedenle metabolizmasını engelleyen ve yarı ömrünü uzatan cilastatin ile birlikte verilir (13). İmipenem, meropenem ve doripenem in vivo yaklaşık 1 saatlik yarı ömürlere sahipken ertapenem, günde bir kez uygulanmaya uygunluk sağlayan yaklaşık 4 saatlik bir yarı ömre sahiptir. İmipenemin diğer karbapenemlere kıyasla doza bağımlı gastrointestinal yan etkileri vardır (14). Ertapenem, imipenem ve meropenem oral olarak emilmez, etkili olması için parenteral uygulama gerektirir (13). Karbapenemler nadiren hepatotoksisiteye neden olabilirler.
2.2. Gram Negatif Basillerde Antimikrobiyal Direnç Mekanizmaları
Bakteriler tarafından β-laktam grubu antibiyotikleri parçalamak için pek çok enzim üretilir. Bu tür enzimler β-laktamazlar olarak bilinir. Bununla birlikte, bakteriler, in vivo kullanım için betalaktam-betalaktamaz inhibitör kombinasyonlarını etkili bir şekilde parçalayamamaktadır.
Antibiyotiklere dirençli bakterilerin neden olduğu enfeksiyonların tedavisi zordur. Bu durum günümüzde gittikçe yaygınlaşmakta ve küresel bir sağlık krizi haline dönüşmektedir. Avrupa İlaç Ajansı’na (EMEA) göre, Avrupa'da antibiyotiğe dirençli bakterilerin neden olduğu ölümlerin yaklaşık üçte ikisi gram negatif bakterilerin neden olduğu enfeksiyonlardan kaynaklanmaktadır (15). Gram negatif bakteriler, ventilatör ilişkili pnömonilerin %45-70’inden, kateter ilişkili kan dolaşımı enfeksiyonlarının %20–30’undan sorumludur. Yoğun bakım ünitelerinde cerrahi alan ve idrar yolu enfeksiyonlarına da neden olur (16).
6
Hastane kökenli enfeksiyon etkeni olan K. pneumoniae, A. baumannii, P. aeruginosa,
Escherichia coli ve Burkholderia cepacia gibi gram negatif patojenlerin, antimikrobiyal
direnç oranları yüksektir (17). Hayatı tehdit edici bu enfeksiyonların tedavisi, uygulanabilir tedavi seçeneklerinin kısıtlı olması nedeniyle giderek zorlaşmaktadır. Enterobacteriaceae familyasının önemli bir üyesi olan K. pneumoniae; yenidoğan ve YBÜ’lerinde özellikle pnömoni ve kan dolaşımı enfeksiyonları gibi hastane enfeksiyonlarının başlıca nedenlerinden birisidir (18). K. pneumoniae’ da, çoklu β-laktamaz enzimlerinin kazanılması; penisilinlere, sefalosporinlere ve karbapenemlere karşı direnç gelişmesine neden olmuştur. Karbapenemler,
K. pneumoniae ve diğer çoklu dirençli gram negatif patojenlerin çoğu için son tedavi
seçeneğidir. Buna karşın, dünya genelinde karbapenemlere karşı artan oranlarda direnç bildirilmektedir. Hatta bazı bölgelerde K. pneumoniae enfeksiyonlarının %50'sinden fazlasında tüm antibiyotik gruplarına karşı direnç bildirilmektedir (19).
KPC üreten K. pneumoniae, bakteriyemi, pnömoni ve idrar yolu enfeksiyonları gibi çok çeşitli enfeksiyonlara neden olur. Bu enfeksiyonlar için, hastanede uzun süre veya yoğun bakım ünitelerinde yatış, immünsupresif veya çoklu antibiyotik tedavisi almak risk faktörleri arasındadır (20). En endişe verici olan, bu organizmaların neden olduğu enfeksiyonlarda tedavi çoklu direnç nedeniyle oldukça kısıtlı ve mortalite oranları yüksektir.
Antimikrobiyal direnç; intrinsik (kromozomal), adaptif veya kazanılmış olabilir. İntrinsik direnç, bir mikroorganizmanın antimikrobiyallerin etkisini sınırlayan doğal özelliklerini ifade eder. Dış zarın geçirgenlik bariyeri ve yapısal olarak eksprese edilen ilaç dışa atım pompaları bunlara örnek verilebilir. Adaptif direnç, bir organizmanın çevresel olaylara yanıt olarak transkriptomlarını hızla değiştirerek birçok stres koşuluna uyum sağlama ve hayatta kalma yeteneğidir. Bu adaptasyonlar organizmaların kısıtlı besin ortamı, antibiyotik stresi gibi koşullarla baş etmelerine yardımcı olur. Kazanılmış direnç; antibiyotiğe duyarlı bir organizmanın direnç genlerini konjugasyon, transformasyon, transdüksiyon gibi çeşitli mekanizmalarla edinmesi veya mutasyonlar sonucunda dirençli hale gelmesiyle ortaya çıkar. Plazmid kaynaklı β-laktam direnç gelişimi, kazanılmış dirence örnek olarak verilebilir (15). Bu mekanizmalar arasındaki sınırlar keskin değildir, örneğin ilaç dışa atım pompaları hem intrinsik hem de kazanılmış dirence katkıda bulunur ve porinler hem adaptif hem de kazanılmış dirençte yer alabilirler.
7 2.2.1. İlacın Hücre İçerisinde Etkin Konsantrasyona Ulaşamaması
2.2.1.1. Antibiyotik Dışa Atımı: İlaç Dışa Atım Pompaları
İlaç dışa atım pompaları, antibiyotiği bakteriyel hücreden dışarı çıkaran proteinlerdir. Böylece hücre içersindeki antibiyotik konsantrasyonunu ölümcül olmayan düzeye düşürür. İlaç dışa atım pompaları, hücrenin ilk savunma hattıdır. Bir mikroorganizma antibiyotik ile karşılaştığında; dışa atım pompa ekspresyonunda geçici upregülasyon olur bu da hücrede subtoksik bir antibiyotik konsantrasyonuna yol açar. Bu nedenle yeni ilaca dirençli mutasyonların seçimi için aktif dışa atım pompası, hem gerekli hem de yeterlidir (21). Bakteriyel dışa atım pompa proteinleri, beş süper aile içerisinde sınıflandırılmıştır. Bunlar MFS, ABC, RND, MATE ve SMR aileleridir. Bu süper aileler hem gram negatif hem gram pozitif bakterilerde görülür ancak RND ailesi büyük çoğunlukla gram negatif bakterilerde saptanır (22). RND aile üyeleri gram negatif bakterilerde klinik olarak anlamlı antimikrobiyal dirence neden olmaları nedeniyle önemlidir (23). Dışa atım pompaları çok sayıda substrat ve antibiyotiği tanır. β- laktam antibiyotikler, RND tipi pompaların substratıdır. Bu kompleksler, gram negatif organizmaların çift zarı üzerinde antibiyotikleri dışarıya atmak için iç zarı, dış zarı ve periplazmik aralığı kapsayan üçlü protein düzenekleridir (24). En iyi bilinen üçlü dışa atım pompa kompleksleri E.coli’nin AcrA–AcrB–TolC ve P. aeruginosa'dan Mex–MexB-OprM taşıyıcılarıdır. Bu komplekslerde AcrB/MexB, ilaçları sitoplazmadan veya periplazmadan atmak için proton hareket kuvvetini kullanan iç zar proteinleridir (25). TolC/OprM, hücre dışına çıkış sağlayan dış zar proteinleridir. AcrA/MexA, iç ve dış zar proteinlerini bağlayan zar füzyon proteinleridir (24, 26). P. Aeruginosa’nın MexAB-OprM RND ailesi, tikarsilinden karbapenemlere kadar bir dizi β laktam antibiyotiğe uyum sağlar. Mex dışa atım sistemlerinin ekspresyonu, P. aeruginosa'daki ertapenem ve aztreonam direnciyle ilişkilendirilmiştir. Enterobacter cloacae'da karbapenem direncine dışa atım pompalarının katkısı da belirlenmiştir (22).
2.2.1.2. Değişmiş dış membran geçirgenliği sayesinde antibiyotik alımında azalma
Gram negatif organizmaların dış membranı, geçirgenlik bariyeri ve birincil savunma hattı olarak işlev görür. β-laktamlar gibi küçük hidrofilik antibiyotikler, gram negatif bakterilerin dış membranlarından porin proteinleri aracılığı ile kolayca gecebilir (27). Porinler, merkezinde hidrofilik bir bölgeye sahip gözenek yapısı oluşturan yapılardır. Bakteriyel porinler substrata spesifik veya difüzyon porinleri gibi spesifik olmayabilir. Yapısal olarak eksprese edilen porinlerin özellikleri, gram negatif bakterilerdeki intrinsik
8
antibiyotik direnci için oldukça önemlidir. Örneğin, P. aeruginosa, antimikrobiyallere karşı
Enterobacteriaceae kökenlerinden çok daha yüksek intrinsik dirence sahiptir. Bu durum P. aeruginosa'nın geçirgenliği yüksek klasik porinler yerine, difüzyon oranları kısmen düşük
olan ve bu nedenle “yavaş” porinler olarak adlandırılan porinleri üretmesinden kaynaklanır (28). P. aeruginosa'nın son derece büyük olan genomu spesifik porinler eksprese etmesine izin verir. Bu porinler küçük besinlerin geçişine izin verirken sefalosporinler gibi antibiyotiklerin geçişine engel olur (29).
Porinler aracılığı ile kazanılmış antibiyotik direnci çeşitli mekanizmalarla oluşur: I. Porinlerin ekspresyonunu azaltan mutasyonlar (örneğin, E.coli'ye β-laktam direnci
sağlayan OmpF kaybı).
II. Büyük çaplı bir porinin daha küçük kanal boyutunda bir porin ile değiştirilmesi (örneğin β-laktam dirençli K. pneumoniae kökenlerinde, OmpK35, daha küçük kanal boyutunda OmpK36 ile değiştirilmesi)
III. Porin işlevini bozan değişikliklere neden olan mutasyonlar (örneğin, N gonorrhoeae'nin PenB porin kanalına negatif yüklü iki amino asidin eklenmesi, penisilin geçişinde belirgin azalmaya neden olur) gibi mekanizmalarla oluşur (28, 30).
Antimikrobiyal direnç gelişiminde, azalmış dış membran geçirgenliği ve dışa atım pompa sistemi arasında önemli bir etkileşim vardır. Bu iki mekanizma birlikte diğer antimikrobiyallere karşı direnç sağlar. Bununla birlikte, antibiyotiklere direnç çoğu zaman tek yönlü olmayıp aynı anda işleyen çoklu moleküler mekanizmalara dayanır. Antimikrobiyallere direnç, azalmış geçirgenlik ve artmış dışa atım pompası, ilaç modifikasyonu ve hedef değişikliği gibi diğer mekanizmaların birlikteliği ile sağlanır.
2.2.1.3. Antibiyotiklerin enzimatik inaktivasyonu: β-laktamaz enzimleri
β-laktam halkası, bu grup antibiyotiklerin ayrılmaz bir parçasıdır. β-laktam antibiyotiklere direnç, temelde β-laktam halkasının amid bağını parçalayarak bu antibiyotikleri inaktive eden enzimler olan β-laktamazların üretilmesiyle oluşur. β-laktamazlar büyük olasılıkla zaman içerisinde doğal antibiyotiklere karşı direnç mekanizması olarak, bakterilerle birlikte evrimleşmişlerdir. Yaygın olarak kullanılan antimikrobiyal tedavinin uyguladığı selektif baskı, bu enzimlerin gelişimini ve yayılmasını hızlandırmış olabilir (6). Modern tıp, β-laktam antibiyotik çeşitliliğini dördüncü kuşak sefalosporinler ve karbapenemlere kadar genişlettikçe, bakteriler bu antibiyotikleri etkisiz hale getirmek ve üstesinden gelmek için, zaman içerisinde çeşitli β-laktamazlar geliştirmiştir. Sürekli artan
9
bakteriyel β-laktamaz enzimleri nedeniyle, β-laktamazların hidrolizden korunabilmesi için klavulanat, sulbaktam ve tazobaktam gibi β-laktamaz inhibitörleri, β-laktam antibiyotiklerle kombinasyon halinde kullanılmaktadır.
β-Laktamazlar, kromozomal genler veya plazmid ve transpozonlar üzerinde bulunan transfer edilebilir genler tarafından kodlanır. Ek olarak, β-laktamaz genleri (bla) sıklıkla çoklu direnç determinantları taşıyan integronlar üzerinde bulunur. Taşınabilir elemanlar tarafından mobilize edilirse, integronlar çoklu bakteri direncinin farklı bakteri türleri arasında daha fazla yayılmasını kolaylaştırabilir (6).
β-Laktamazlar amino asit yapılarına göre ilk olarak Ambler tarafından önerilen şekilde A ile D arasındaki dört moleküler sınıfa ayrılır. Alternatif olarak, Bush-Jacoby-Medeiros sistemi, enzimleri substrat profillerine göre sınıflandırır. Klavulanik asit gibi β-laktamaz inhibitör duyarlılığı, betalaktamazları birkaç fonksiyonel gruba ayırır. Sınıf A, C ve D β-laktamazların β-laktam halkasının aktif bölgesinde bir serin amino asidi vardır. B sınıfı enzimlerin aktif bölgesinde ise çinko (Zn2+) bulunmaktadır (6). Betalaktamazların sınıflandırılması Tablo 1.’ de gösterilmiştir.
10 Tablo 1. β-Laktamazların Sınıflandırması (31)
Bush-Jacoby grup (2009) Bush-Jacoby Medeiros grup (1995) Moleküler sınıflama (Alt sınıf) Ayırt edici substrat İnhibe edici Temsilci enzimler Klavulanik asit veya tazobaktam EDTA
1 1 C Sefalosporinler Hayır Hayır
E. coli AmpC, P99, ACT-1, CMY-2,
FOX-1, MIR-1
1e İçermez C Sefalosporinler Hayır Hayır GC1, CMY-37
2a 2a A Penisilinler Evet Hayır PC1
2b 2b A
Penisilinler ve erken sefalosporinler
Evet Hayır 1, TEM-2, SHV-1 2be 2be A Genişletilmiş spektrumlu sefalosporinler, Monobaktam Evet Hayır TEM-3, SHV-2, CTX-M-15, PER-1, VEB-1
2br 2br A Penisilinler Hayır Hayır TEM-30,
SHV-10 2ber İçermez A Genişletilmiş spektrumlu sefalosporinler, Monobaktam
Hayır Hayır TEM-50
2c 2c A Karbenisilin Evet Hayır PSE-1,
CARB-3
2ce İçermez A Karbenisilin,
sefepim Evet Hayır RTG-4
2d 2d D Kloksasilin Değişken Hayır 1,
OXA-10
2de İçermez D
Genişletilmiş spektrumlu sefalosporinler
Değişken Hayır OXA-11, OXA-15
2df İçermez D Karbapenemler Değişken Hayır OXA-23,
OXA-48
2e 2e A
Genişletilmiş spektrumlu sefalosporinler
Evet Hayır CepA
2f 2f A Karbapenemler Değişken Hayır KPC-2, IMI-1,
SME-1
3a 3
B (B1)
Karbapenemler Hayır Evet
IMP-1, VIM-1, CcrA,
IND-1
B (B3) L1, CAU-1,
GOB1, FEZ-1
3b 3 B (B2) Karbapenemler Hayır Evet CphA, Sfh-1
11 2.3.Karbapenem Direnç Gelişimi
Karbapenemlere karşı direnç gelişimi, intrinsik veya kazanılmış direnç mekanizmalarıyla ayrı ayrı veya birlikte olmasından kaynaklanabilir. Karbapenem direnci;
I. Karbapenemleri hidrolize eden β-laktamaz üretimi (karbapenemaz),
II. Dış membran proteinlerinin ekspresyonunda azalma nedeniyle porin kaybı sonucunda membran gecirgenliğinin azalması,
III. Dışa atım pompaları sayesinde ilacın hücre dışına atılması,
IV. Değişime uğramış veya düşük afiniteli PBP üretimi gibi mekanizmalarla gelişir (6).
Gram pozitif kok ve gram negatif basillerin karbapenem direnç gelişim özellikleri birbirinden farklılık gösterir. Gram pozitif koklarda, karbapenem direnci tipik olarak PBP amino asit dizilerindeki yer değişimine ya da yeni kazanılmış bir PBP üretimine bağlıdır. PBP değişimine bağlı direnç, gram negatif basillerde nadir görülür. Gram negatif basillerde, karbapenem direnci genellikle karbapenemaz üretimi, dışa atım pompaları, porin kaybı gibi bir veya birden fazla mekanizma aracılığı ile gelişir (6)
2.3.1.Karbapenemazlar
Karbapenemazlar, yalnızca karbapenemleri değil aynı zamanda geniş spektrumlu penisilinler, oksiimino-sefalosporinler ve sefamisinleri de hidrolize edebilir. Bu nedenle oldukça geniş direnç spektrumununa sahiptirler (6).
2.3.1.1.Karbapenemazların Moleküler Sınıflandırılması
Enterobacteriaceae’ya ait karbapenemazlar Ambler A, B ve D grubu β-laktamaz
sınıfında yer alır. Bu sınıflar nozokomiyal patojenler arasında klinik olarak en önemli gruplardır. Sınıf A ve D β-laktamazların tümü aktif bölgede bir serin rezidüsü taşır, bu nedenle serin betalaktamazlar olarak adlandırılır. B sınıfı β laktamazlar aktif bölgelerinde Zn2+ iyonu bulundururlar bu yüzden metallobetalaktamazlar olarak adlandırılır (6).
2.3.1.1.1. A Sınıfı Karbapenemazlar
Moleküler A sınıfı serin karbapenemazlar nadir olarak görülür ve genellikle
Enterobacteriaceae izolatlarında rastlanırlar. Bush ve arkadaşlarının önerdiği sınıflandırma
12
kodlayan genler, kromozom veya plazmidler üzerinde bulunur. Bu sınıfta yer alan enzimler; NMC, SME, IMI kromozomal olarak kodlanırken; KPC (KPC-2 ila KPC-13), GES türevleri (GES-1 ila GES-20) plazmid kodludur (10, 32). Ancak bunların hepsi karbapenemleri aktif olarak hidroliz eder ve kısmen klavulanik asit tarafından inhibe edilir (33, 34).
2.3.1.1.1.1.Plazmid Kodlanmış Enzimler: KPC ve GES 2.3.1.1.1.1.1.KPC
A sınıfı serin karbapenemazlar arasında en yaygın olanları KPC enzimleridir. KPC karbapenemazları, iki karakteristik özellikleri ile diğer fonksiyonel grup 2f enzimlerinden ayrılır. İlk olarak, KPC enzimleri transfer edilebilir plazmidlerde bulunur. İkinci olarak substrat hidroliz spektrumu, sefotaksim gibi sefalosporinleri de içerir (35) . K. pneumoniae, KPC'leri taşıyan en yaygın bakteri türü olmaya devam etse de, bu enzimler diğer gram negatif basillerde de tanımlanmıştır (36). BlaKPC, Kolombiya'daki hastalardan elde edilen P. aeruginosa kökenlerinde ve Porto Riko'daki hastalardan elde edilen Acinetobacter türlerinde
de saptanmıştır (37, 38). KPC enzimi salgıladığı belirlenen bakteri türleri Tablo 2.’de gösterilmiştir.
Tablo 2. KPC saptanan türler (36)
Enterobacteriaceae Enterobacteriaceae dışı
Citrobacter freundii Pseudomonas aeruginosa
Escherichia coli Pseudomonas putida
Enterobacter aerogenes Acinetobacter spp
Enterobacter cloacae Enterobacter gergoviae Klebsiella pneumoniae Klebsiella oxytoca Proteus mirabilis Salmonella enterica Serratia marcescens
13
KPC ailesinin bir üyesi olan KPC-1 1996 yılında Kuzey Carolina'da ICARE sürveyans projesi sırasında bir K. pneumoniae klinik izolatında ilk kez saptandı (39). KPC-2 ise ilk olarak 2003 yılında KPC-1'deki bir nokta mutasyonunun sonucu olarak tanımlanmış ve 1998'den 1999'a kadar Baltimor’da dört kökende ortaya çıkmıştır. Bu dört kökende KPC-2 üreten genin, transfer edilebilir bir plazmid üzerinde yer aldığı ve tüm izolatlarda, imipenem'e karşı duyarlılığın azaldığı görülürken, hiçbirinin CLSI kriterlerine göre dirençli olmadığı gözlenmiştir. Daha sonra KPC-2, Maryland’ın başka bir bölgesinde Salmonella enterica'daki bir plazmidde gösterilmiştir (35).
New York’da 2002 yılı sonunda görülen ilk hastane salgınına kadarki dönemde, K.
pneumoniae'da karbapenem direnci nadir olarak görülmekteydi (40). İlk salgınlarla birlikte,
bu bakterilerde penisilin, sefalosporin, monobaktam ve karbapenemler dahil tüm beta-laktamlara karşı duyarlılıkta azalma veya direnç belirlenmesi büyük endişe yarattı. Dahası, 2003-2004 yılları arasında Brooklyn hastanelerinde soyutlanan 95 KPC enzimi üreten kökenin in vitro çalışmaları, bu kökenlerin yarısının aminoglikozitlere ve yarısından fazlasının da flourokinolonlara dirençli olduğunu ortaya koydu (36). New York'taki ilk sporadik salgınların ardından, KPC üreten bakteri, New York ve New Jersey bölgelerindeki birçok hastanede endemik haleeldi. CDC'ye bildirilen nozokomiyal enfeksiyonlara ilişkin veriler, K. pneumoniae’daki karbapenem direnç oranının, 2000 yılından 2007 yılına kadar yüzde birden yüzde sekize yükseldiğini göstermiştir (41). ABD dışında KPC üreten ilk köken 2005 yılında Fransa'dan bildirildi. Bu kökenin de yakın zamanda New York'ta hastanede yatmış olan bir hastadan soyutlandığı belirlendi (42). ABD dışındaki ilk salgın ise İsrail’den raporlandı. İsrail (ST258), Kore (ST258) ve Çin (ST11), yüksek oranda KPC-2 ve KPC-3 prevalansına sahip olan ülkelerdir. Ayrıca KPC üreten K. pneumoniae ile ilgili çok merkezli yayınlar bulunmaktadır. Bu yayınlarda; Polonya ve Fransa’dan KPC-2 (ST258), Singapur ve Tayvan’dan KPC-2 (ST11) bildirilmiştir. Kıbrıs ve özellikle İtalya’dan ise KPC-2 ve KPC-3 (ST258 / 512 ve diğer ST'ler) bildirilmiştir. Günümüzde Akdeniz ülkelerinde birçok sağlık kurumunda KPC üreten K. pneumoniae suşları baskındır (43, 44). Son zamanlarda Yunanistan'da KPC (ST258) prevalansında çarpıcı bir yükseliş eğilimi gözlenmektedir (32). KPC’ler İsrail ve Yunanistan’da endemik hale gelmiştir (36).
KPC üreten bakterilerin moleküler epidemiyolojisine daha yakından bakıldığında,
blaKPC yayılımından, birkaç klonun sorumlu olduğu ortaya çıkmıştır. KPC-2 taşıyan bir K. pneumoniae klonu, ST-258, geniş bir coğrafyada blaKPC-2 yayılımına neden olmuştur.
14
transpozon P. aeruginosa plazmidi olan pCOL-1'de de gösterilmiştir. Bu durum transpozonun farklı gram negatif türler arasında aktarılma olasılığını düşündürmüştür (45).
1996-2008 yılları arasında ABD, İsrail ve Hindistan'dan CDC'ye gönderilen KPC üreten K. pneumoniae kökenleri incelenmiştir. CDC veritabanındaki suşların yaklaşık %70'inde ve İsrail’den gelen kökenlerin tümünde ST258’in baskın suş olduğu gözlenmiştir. Yedi farklı enzim tipi bildirilmiş olmasına rağmen ST258 suşlarının çoğunun KPC-3'ü, ST258 dışı suşların çoğunun ise KPC-2'yi ürettiği belirlenmiştir (36). Türkiye’den bildirilen ilk KPC üreten K. pneumoniae kökeni Romanya’dan transfer edilen bir hastada bildirilmiştir (46). Daha sonra Türkiye’den KPC üreten K. pneumoniae ve E. coli izolatları raporlanmıştır (47, 48).
BlaKPC’nin transfer edilebilir plazmidlerde bulunması enfeksiyon kontrolünü zorlaştırır
ve kritik hastalarda hayati tehdit oluşturur. K. pneumoniae'nın blaKPC’yi çevreleyen plazmidler
üzerinde yapılan sekans analizinde; transpozaz (tnpA), resolvaz (tnpR) ve iki bağımsız insersiyon dizisinin mobil Tn4401 üzerinde değişmez bileşenler olduğu ortaya koyulmuştur (49). Yakın zamanda yapılan in vitro konjugasyon çalışmalarında, blaKPC-2'nin, Escherichia coli'deki IS26 ve transfer operonu (locus tra-trb) yardımı ile transfer edilebileceği
gösterilmiştir, ancak bu mekanizma K. pneumoniae'da bulunmamıştır (50). Plazmidin horizontal transferi, blaKPC’nin farklı Enterobacteriaceae cinsleri arasında yayılımına katkıda
bulunmuştur. Bu nedenle BlaKPC-2’ninçapraz tür aktarımı endişe vericidir (51).
KPC enzimleri, A sınıfı β-laktamazların korunmuş aktif bölge motifleri olan S-X-X-K, S-D-N ve K-T-G'ye sahiptir. Bu enzimler, SME karbapenemazlara %45 benzerlik oranı ile en yakın amino asit dizisine sahip karbapenemazlardır. KPC’ler, SME ve NMC/IMI enzimleri için tarif edilen, disülfid bağına sahip, korunmuş C69 ve C238 kalıntılarına sahiptir (35). KPC enzimlerinin yapısal analizi, bu enzimlerde katalitik serinin, diğer A sınıfı β-laktamazlardan farklı olarak daha yüzeysel bir pozisyonda olduğunu göstermiştir. Bunun sonucunda KPC enzimleri karbapenmleri diğerlerine göre daha kolay yıkar (52).
Çalışmalarda, KPC salgılayan K. pneumoniae izolatlarının AmpC sefalosporinaz (blaDHA ve blaCMY), genişletilmiş spektrumlu β-laktamaz (blaCTX-M) ve MβL'leri kodlayan
plazmid kökenli genleri de birlikte taşıdığını göstermiştir (53). Ayrıca, blaKPC taşıyan
plazmidler kinolon-direnci (qnrA, qnrB) ve aminoglikozit modifiye edici genetik determinantları (ArmA ve RmtB) da bulundurabilir (54). Bunun sonucunda, KPC üreten gram negatif bakteriler çoğunlukla antimikrobiyal seçenekleri kısıtlı, çoklu dirençli (MDR) fenotipler sergiler. KPC salgılayan K. pneumoniae kökenlerinde, karbapenemlerin hücre içine
15
girişini sağlayan OmpK36 porin eksikliğine bağlı olarak, direnç düzeyinin arttığı bazı çalışmalar ile gösterilmiştir (55, 56).
2.3.1.1.1.1.2.GES
GES/IBC ailesi ilk kez 2000 yılında tarif edilen nadir görülen bir ailedir. IBC-1 Yunanistan'da bir E. cloacae kökeninden raporlanmıştır. GES-1 ise Fransız Guyanası'ndan bir K. pneumoniae’da bildirilmiştir. Bu enzimler birbirinden sadece iki amino asidin yer değişikliği ile farklılık gösterir. A sınıfı karbapenemazlarda olan Ambler pozisyonları 69 ve 238'deki sistein kalıntıları ve aktif bölge motifleri GES karbapenemazlarda da bulunur. GES/IBC ailesi; KPC-2 ile %36 , SME-1 ile %35 ve NMC ile %31 oranında amino asit dizi benzerliğine sahiptir. Bu durum uzaktan ilişkili olduklarını göstermektedir (35). GES enzimlerini kodlayan genler, plazmidler üzerindeki integronlarda bulunur. Daha önceleri GES enzimleri, ESBL enzimleri içerisinde yer almaktaydı. Ancak 2001 yılında GES-2 üreten P.
aeruginosa'da bu enzimin imipenemi hidrolize edebildiği belirlendikten sonra
karbapenemazlar arasına alındı. GES enzimlerinin tüm varyantları karbapenemaz enzimleriyle uzaktan ilişkilidir (32).
GES enzimleri nadir olmasına rağmen, Yunanistan, Fransa, Portekiz, Güney Afrika, Fransız Guyana’sı, Brezilya, Arjantin, Kore ve Japonya'dan sporadik olgular raporlanmıştır. Ancak, GES-2'yi eksprese eden P. aeruginosa suşları ile Güney Afrika’dan sekiz hastada küçük bir nozokomiyal salgın, Kore'den ise altı hastada GES-5 üreten K. pneumoniae enfeksiyonları bildirilmiştir (35, 57).
2.3.1.1.1.2. Kromozomal Kodlanmış Enzimler: SME, NMC ve IMI
NMC/IMI ve SME enzimlerini kodlayan genler kromozomaldir. Bu enzimler imipenemi meropenemden daha kolay hidrolize eder. Karbapenem direnç düzeyini; nmcR, ImiR ve smeR gibi düzenleyici genlerin ekspresyonu belirler. SME-1 ilk kez 1982 yılında İngiltere'den iki S. marcescens izolatında bildirildi. Amerika’dan da sporadik SME-2 ve SME-3 raporları bildirilmiştir. SME-2 ve SME-3 yapısal olarak SME-1 ile büyük oranda benzerlik gösterir (32).
IMI ve NMC enzimleri ABD, Fransa ve Arjantin'de, E. Cloacae kökenlerinde nadir olarak bildirilmiştir. NMC enzimleri ile IMI-1 arasında %97, SME-1 arasında ise %70 oranında amino asit benzerliği mevcuttur. Bu karbapenemazların hepsi, A sınıfı
β-16
laktamazlarda gözlendiği gibi S-X-X-K, S-D-N ve K-T-G korunmuş aktif alan motiflerine sahiptir. Ayrıca diğer A sınıfı β-laktamazlara benzer şekilde bunlarda da disülfit köprüsü oluşturan 69. ve 238. pozisyonlarında sistein kalıntıları vardır. Bu enzimlere ait genler kromozomaldir ve mobil elemanlarla ilişkisi gösterilememiştir. Nadir saptanmaları buna bağlı olabilir. Ancak, yakın zamanda yapılan araştırmalarda, plazmid kökenli IMI-2 enzimleri taşıyan ABD’den Enterobacter asburiae ve Çin'den ise E. cloacae bildirilmiştir (35). IMI-2’nin plazmid ile ilişkisinin bildirilmesi yayılım açısından daha dikkatli izlemini gerektirir.
2.3.1.1.2. B sınıfı karbapenemazlar
Moleküler B sınıfı MβL'ler, karbapenemler dahil tüm β-laktamları hidrolize edebilir. Sulbaktam, klavulanat ve tazobaktam gibi β-laktamaz inhibitörlerine direnç gösterir. Ancak aztreonam'a duyarlıdırlar ve EDTA gibi metalik iyon şelatörleri tarafından inhibe edilirler. Etki mekanizması temelde β-laktam substratların, enzimin etki alanında bulunan Zn+2 iyonu ile etkileşimine bağlıdır. Genişletilmiş spektrumlu sefalosporin veya karbapenemlerin sık kullanımı, MβL’ların seleksiyonuna ve yaygınlaşmasına yol açmıştır (35).
Kromozomal yerleşimli MβL enzimleri ilk kez, Bacillus cereus, Aeromonas spp.,
Elizabethkingia meningoseptica ve Stenotrophomonas maltophilia gibi çevresel, fırsatçı
patojenlerde bildirilmiştir. Kromozomal MβL genleri kolayca transfer edilemez. Bununla birlikte, S. maltophilia ve E. meningoseptica ile nozokomiyal salgınlar rapor edilmiştir (32, 58, 59).
2002'den önce, Japonya'da gram negatif bakterilerde MβL enzimlerinin genel prevalansı %0.5 olmakla birlikte S. marcescens'te %3.0 ve imipenem dirençli P. aeruginosa'da % 2.6 oranındadır (60). Ancak daha sonraları plazmid aracılı MβL'ların oranında hızlı bir artış olmuştur. Dünya genelinde en sık rastlanan MβL enzimleri arasında IMP, VIM ve NDM yer alır. GIM, SPM ve SIM ise endemik olarak bulunmaktadır. NDM, GIM ve SPM dışındaki MBL genleri, genellikle plazmid üzerindeyken yayılmalarını kolaylaştıran integron kasetlerinde yer alırlar (32).
Proteinlerin analizine dayalı ilk sınıflandırma, MBL’lerin serin β-laktamazlardan farklı olduğunu gösterdi. MBL’lerin β-laktam hidrolizi için Zn2+’ya gereksinim göstermesi yanısıra
klavulanik asit ve tazobaktam tarafından inhibe edilememesi, en önemli ayırt edici özellikleridir. Metalo-β-laktamazlar, β-laktamaz aktivitesinin kolorimetrik göstergesi olan nitrosefini kolayca hidrolize edemez (35).
17
İlk kez laktamazlar Bacillus cereus'ta gösterilmiş ve BCII metalo-β-laktamazı uzun yıllar prototip olarak kabul edilmiştir. En yaygın metalo-β-laktamaz aileleri olan VIM, IMP, GIM ve SIM enzimlerinin gen kasetleri çeşitli integronlarda yer alır. Bu integonların plazmid veya transpozonlar gibi mobil elemanlarla etkileşimi bakteriler arası transferi kolaylaşır (35).
2.3.1.1.2.1.IMP
IMP enzimleri ilk olarak 1991'de klinik örneklerdeki P. aeruginosa plazmidlerinde saptandı ve daha sonra 1995'te Japonya'da S. marcescens'te gösterildi (61, 62). Bu enzim penisilin, genişletilmiş spektrumlu sefalosporinleri ve imipenemi hidrolize ederken aztreonama etkili değildir. Sınıf 1 integron gen kasetinde bulunan IMP-2 genleri ilk olarak 2000 yılında İtalya'da Acinetobacter baumannii kökenlerinde belirlendi (63). Mobil integonlar, blaIMP ve qacE1, sul1 direnç genlerinin ekspresyonunu kolaylaştıran integraz,
rekombinaz ve promotör gibi önemli intrinsik genetik belirteçleri içerir. QacE1ve sul1 sülfonamidlere karşı, blaIMP ise kuarterner amonyum bileşiklerine karşı direnç kodlayan
genlerdir. Geçmişten günümüze değin IMP ailesi tüm dünyada gözlenmiş ve son olarak ABD ve Avustralya'ya yayılmıştır (64). Bununla birlikte IMP geni, endemik olarak sadece Tayvan'da (blaIMP-8) ve Japonya'da (blaIMP-1) bildirilmiştir (65, 66).
2.3.1.1.2.2.VIM
Sık rastlanan metallo-β-laktamaz ailelelerinden bir diğeri VIM enzimleridir. VIM enzimlerini kodlayan genler de BlaIMP’ye benzer şekilde, K. pneumoniae üzerinde plazmid
ilişkili sınıf 1 integronlarda yer alır (67). VIM genlerini taşıyan integronlar, özellikle
Enterobacteriaceae kökenlerinin plazmid veya kromozomlarına yüksek oranda ve kolayca
konjügasyon yeteneğine sahiptir. Böylece, bu özellik blaVIM’in gram negatif bakterilerde, aynı
cinsdeki farklı türler veya aynı türdeki farklı klonlar arasında hızlı bir şekilde yayılmasını sağlar (68). VIM ve IMP aileleri, plazmid üzerinde taşınmaları ve integronla ilişkili olmaları nedeniyle bazı benzerliklere sahiptir. Her ikisi de, monobaktamlar dışındaki tüm β-laktamları hidrolize eder ve tüm β-laktam inhibitörlerine karşı duyarlıdır (69). BlaIMP ve blaVIM
arasındaki konjugasyon etkinliğindeki fark, dünya çapında blaVIM taşıyan gram negatif
bakterilerin daha yüksek oranda olmasını açıklar. Ayrıca, çalışmalarda, blaVIM içeren
18
direnç genlerinin de taşınabildiği gösterilmiştir (32). Bu nedenle, bazı blaVIM taşıyan K. pneumoniae izolatları çoklu direnç fenotipleri gösterir.
VIM enzimleri ilk kez 1996 yılında Fransa’dan 2) ve 1997’de İtalya’dan (VIM-1) P. aeruginosa kökenlerinde bildirilmiştir. (70, 7(VIM-1). Günümüze kadar VIM-1, VIM-2 ve VIM-5 Avrupa ülkelerinden, VIM-5 Türkiye’den, VIM- 3 Tayvan’dan, VIM-2 ise Tunus, ABD, Kore, Japonya ve Güney Amerika ülkelerinden bildirilmiştir. VIM enzim varyantlarını kodlayan genler Pseudomonas spp, A. Baumannii K. pneumoniae, E. coli, E. cloacae ve P.
mirabilis kökenlerinde saptanmıştır (67, 72-75). Bir sürveyans çalışması, blaVIM-1 taşıyan
gram negatif bakteri enfeksiyonlarının 2010 yılında Yunanistan'da endemik düzeye ulaştığını bildirmiştir. Bu durum sonucunda Yunanistan blaVIM, blaKPC-2 (ST258) ve blaNDM-1 aktarımı
için potansiyel bir merkez haline gelmiştir (76). Tayvan'da, 2000-2002 döneminde antimikrobiyal direnç sürveyans çalışması, MβL genlerinin prevalansının %17-36 arasında olduğunu göstermiştir. Çalışmada VIM-3, karbapenem dirençli P. aeruginosa izolatlarında % 88 oranıyla en yaygın saptanan MβL enzimidir (77). Tayvan'da 2000-2010 yılları arasında yapılan sürveyans çalışması, P. aeruginosa’larda karbapenem direncinde anlamlı bir artış olduğunu göstermiştir. Kökenlerde karbapenem direncinin %18,3 oranında melatallo-β-laktamaz enzimlerinden kaynaklandığı belirlenmiş, bunların tamamının VIM enzimleri olduğu gösterilmiştir (78).
2.3.1.1.2.3.NDM
NDM enzimleri ilk olarak 2008 yılında, New Dehli’de hospitalize edildikten sonra İsviçre’ye transfer edilen bir olgunun idrarından soyutlanan K. pneumoniae’da saptandı (79). NDM genleri sıklıkla K. pneumoniae ve E. coli kökenlerinde saptanmasına rağmen A.
baumannii ve P. aeruginosa suşlarında da gösterilmiştir (10). BlaNDM farklı mekanizmalarla
yayılabilmektedir. Örneğin, blaNDM-1’nin aynı hastada K. pneumoniae ve E. coli gibi farklı
cinsler arasında in vivo olarak transfer edilebildiği gösterilmiştir (79). Ayrıca, Çin'de yapılan bir araştırmada blaNDM-1 taşıyan E. coli suşlarının genetik ortamı incelendiğinde; NDM-1
geninin IS3000, ISAba125, IS26 ve bir transpozas ile çevrili olduğu gösterilmiştir. Bu durum araştırıcılar tarafından E. coli ve çevresel Acinetobacter türleri arasında mobil transpozonlar aracılığı ile yatay blaNDM-1 transferi olduğu şeklinde yorumlanmıştır (80). Avustralya’da hiç
yurtdışına çıkmamış bir çocukta ev içi temaslar sırasında blaNDM-1 alellerinin yayılabildiği
gözlenmiştir (81). New Delhi'de, içme sularında blaNDM-1 bulunduran çeşitli bakteriler
19
(82). Sonuç olarak, NDM-1 genlerinin yayılm mekanizmaları karmaşıktır, bu nedenle enfeksiyon kontrolü güçtür.
BlaVIM ve blaIMP'den farklı olarak, blaNDM integron aracılı yayılım göstermez. BlaNDM’nin plazmid aracılı yayılımında olası düzelenleyici mekanizmalardan
bahsedilmektedir. Bunlardan birisi blaNDM taşıyan kasetlerin, plazmid integronlarında yeniden
yerleştirilmesidir. Diğeri ise IS26, ISEc33 veya ISSen4 gibi bazı insersiyon elemanlarının yardımıyla blaNDM ile ilişkili segmentlerin rekombinasyon veya transpozisyonudur (35). Son
zamanlarda klinik örneklerden izole edilen A. baumannii kökenlerinde , blaNDM-1’in Tn125
aracılığı ile yayılabildiği gösterilmiştir (83). BlaNDM-1 taşıyan plazmidler; D sınıfı oksasilinaz,
plazmid aracılı AmpC sefalosporinaz, CTX-M gibi ESBL enzimlerini kodlayan genler yanısıra, makrolid, sulfametoksazol, rifampisin ve aminoglikozidlere karşı dirençten sorumlu diğer genleri de içerir (33, 84). Bunun sonucunda blaNDM-1 içeren gram negatif bakteriler
genellikle çoklu direnç profili göstermektedir.
NDM-1 üreten enterik gram negatif bakteriler Orta ve Güney Amerika dışında çoğu kıtadan bildirilmiştir (33). Avustralya'da son zamanlarda blaNDM bulunduran Enterobacteriaceae kökenlerinin bildirimleri artmaktadır (85-87). Ayrıca, Birleşik Arap
Emirlikleri ve Balkan ülkeleri, blaNDM-1'in Hindistan’dan sonraki ikinci rezervuarı haline
gelmiştir (77, 88, 89). Hindistandaki salgınlarla ilişkisi olmayan blaNDM bildirimlerinin farklı
ülkelerde de artması dikkat çekmektedir.
2.3.1.1.2.4.SPM, GIM ve SIM
SPM-1 ilk kez 2004 yılında Brezilya'nın Sao Paulo kentinde P. aeruginosa
kökenlerinde tanımlandı. BlaSPM-1'in genetik analizi, bu genlerin integronun bir parçası
olmadığını ancak transpozonlarda olduğunu gösterdi. Bu nedenle, blaSPM-1 nozokomiyal
salgınlara neden olma potansiyeline sahiptir (90). GIM-1 enzimi ise 2002 yılında Almanya'da
P. aeruginosa'nın taşınmaz plazmidlerinde bulundu, daha yakın tarihlerde
Enterobacteriaceae’da da saptandı (91). BlaSIM-1 genleri, Kore'de A. baumannii kökenlerinde
20 2.3.1.1.3.D sınıfı karbapenemazlar: OXA β-laktamazlar
Tarif edilen ilk OXA enzimleri, benzilpenisiline göre oksasilini daha yüksek oranda hidrolize etmesi nedeniyle oksasilinazlar veya OXA β-laktamazlar olarak adlandırılmıştır. Bu genelleme artık günümüzde geçerli değildir (93). İşlevsel olarak, kloksasilin ve oksasilin'i hidrolize edebilen penisilinazlardır. Karakteristik olarak klavulanat veya EDTA tarafından inhibe edilmezler (32). D sınıfı β-laktamazlar çok geniş bir aile olup, şu anda 400'den fazla varyantı bulunmaktadır. Bu varyantlar arasında bir kısmının karbapenemaz aktivitesi vardır (94).
Karbapenemaz aktivitesine sahip ilk OXA β-laktamaz, 1985 yılında Edinburgh, İskoçya'daki bir hastanın kanından soyutlanan çoklu dirençli A. baumannii suşunda belirlenmiştir (95). İlk kez 1993 yılında Paton ve arkadaşları tarafından ARI 1 (Acinetobacter resistant to imipenem) enzimi olarak isimlendirilmiştir. ARI 1 enzimi, gen dizilimi sonrası Ambler D sınıfı OXA-23 enzimi olarak sınıflandırılmıştır (96). OXA karbapenemazların büyük çoğunluğu, A. baumannii kökenlerinde görülmektedir. D sınıfı kazanılmış karbapenemaz genleri taşıyan A. Baumannii kökenleri, %30’un üzerinde mortalite oranıyla çok sayıda nozokomiyal salgına neden olmaktadır. Brezilya, İngiltere, Kore ve Tahiti’deki karbapenem dirençli Acinetobacter salgınlarında, OXA-23 karbapenemazlar bildirilmiştir. İki amino asit ile birbirinden ayrılan OXA-24 ve OXA-40, İspanya ve Portekiz'deki hastanelerde klonal Acinetobacter salgınlarına neden olmuştur. OXA-23 ve OXA-58 karbapenemaz üreten
Acinetobacter suşları, 2003 ve 2005 yılları arasında Irak ve Afganistan'da görev yapan askeri
ve sivil personelde enfeksiyonlara neden olmuştur (97).
Amino asit sekans çalışmalarında, D sınıfı karbapenemazlar OXA-23, OXA-24/40, 48, 51, 58, 134a, 143, 211, 213, 214, OXA-229 ve OXA-235 olarak 12 alt gruba sınıflandırılmıştır. Özgün bir sekans hatası nedeniyle OXA-24 günümüzde OXA-40 olarak değiştirilmiş ve böylece OXA-24/40 olarak yeniden tanımlanmıştır (94). OXA-23 enzimi ilk olarak 1985'te İngiltere'de A. baumannii’de bildirilmiştir. Günümüze kadar, blaOXA-23 için 18 varyant tanımlanmıştır (95). BlaOXA-23,
kromozom veya plazmidlerde yer alabilir. OXA-24/40, İspanya'da 1997 yılında A.
baumannii kökenlerinde tanımlanmış ve altı varyant bildirilmiştir (98). OXA-48, ilk kez
Türkiye’de 2001 yılında bir K. pneumoniae kökeninde saptanmıştır. Bu alt grubun günümüze kadar 10 varyantı belirlenmiştir (99). D sınıfı karbapenemazlar arasında imipenem hidrolizi en güçlü olan OXA-48’dir (35). OXA-51, OXA tipi β-laktamazlar arasındaki en büyük alt gruptur. Bu enzim ilk olarak 1996'da Arjantin'de yine bir A. baumannii kökeninde tanımlanmış olup günümüze dek bildirilen varyant sayısı 94’e ulaşmıştır. A. baumannii
21
suşları genellikle blaOXA-51’i doğal olarak taşır. Fransa'da 2003'te çoklu dirençli A. baumannii suşlarında ilk kez gösterilen OXA-58’in üç varyantı bulunmuştur. OXA-134a,
farklı bir tür olan Acinetobacter lwoffii kökeninde tanımlanmış ve altı varyantı mevcuttur. Benzer şekilde yine aynı tür olan A. lwoffii kökeninde ilk kez 2004 yılında Brezilya'da gösterilen OXA-143’ündört varyantı tanımlanmıştır (100). Son zamanlarda yapılan çalışmalar ile daha önce bahsedilen 12 alt grubun dışında yeni alt gruplar tanımlanmıştır. Bunlar;
Acinetobacter johnsonii'den OXA-211, Acinetobacter calcoaceticus'tan OXA-213,
Acinetobacter haemolyticus'dan OXA-214, Acinetobacter bereziniae’den OXA-229, ve Acinetobacter schindleri kaynaklı OXA-235 enzim alt gruplarıdır (94, 100).
D sınıfı β-laktamazların çok geniş bir aile olması literatürde bu sınıfın tanımlanmasında karışıklığa neden olabilmektedir. Farklı amino asit dizilimine sahip çok sayıda alt gruba rağmen D Sınıfı β-laktamazlarda korunmuş amino asit kalıntıları ve motifleri mevcuttur.D sınıfı karbapenemazların dizi analizi; katalitik serin ve lizin kalıntılarının 70 ila 73. pozisyondaki iyi korunan Ser-Thr-Phe-Lys tetradında bulunduğunu ortaya koymuştur. Neredeyse özdeş bir amino asit motifi, A sınıfı β- laktamazlarda da bulunmaktadır (35, 93). Tyr-Gly-Asn 144-146. pozisyonda yer alan ve korunan ikinci bir motiftir. Yine büyük ölçüde korunan bir başka bölge de 216-218. pozisyondaki Lys-Thr-Gly (veya Lys-Ser-Gly) motifidir. Diğer korunan kalıntılar arasında Ser118, Gly131, Trp164, Leu189, Trp234 ve Gly237 bulunmaktadır (93).
2.3.1.1.3.1. OXA-48:
OXA-48 enzimi ilk kez 2001 yılında İstanbul’da hastanede yatmakta olan bir olgunun idrarından soyutlanan K. pneumoniae’da tanımlandı (99, 101). OXA-48 enzimi, OXA-54, OXA-181 ile birlikte Shewanella oneidensis, K. pneumoniae ve diğer enterobakterilerde gösterildi. Shewanella oneidensis progenitör blaOXA-48 benzeri genlerin doğal bir taşıyıcısıdır.
KPC enzimlerinden farklı olarak OXA-48’in karbapenemi hidrolizi sırasında, karbapenemdeki α-hidroksietil grubunun aktif bölge içerisinde dönmesi gerekmektedir. Bu durum KPC ile karşılaştırıldığında, OXA-48’de imipenem hidrolizinin daha zayıf oluşunu açıklar. Aslında, OXA-48'in imipenem'e karşı in vitro hidrolitik etkinliği Acinetobacter
spp'deki diğer oksasilinazlardan yaklaşık on kat daha yüksektir (32). BlaOXA-48 transfer
edilebilir transpozon Tn1999 (veya Tn1999.2) ile plazmitler üzerinde taşınır, orta veya yüksek düzeyde yayılma potansiyeline sahiptir (35). BlaOXA-48 benzeri genlere sahip enterik gram
22
birlikte taşıyabilir. Böylece aztreonam, genişletilmiş spektrumlu sefalosporinler ve karbapenem ajanlara karşı direnç gösterebilir (32).
Plazmid üzerindeki spesifik konumlarından dolayı, blaOXA-48 dünya çapında yayılması
şaşırtıcı değildir. BlaOXA-48 taşıyan K. pneumoniae izolatları ilk olarak Türkiye'de
tanımlandıktan sonra Orta Doğu, Kuzey Afrika ve İngiltere, Belçika, Fransa, Almanya, Hollanda gibi Batı Avrupa ülkelerinde saptandı. Hindistan, Senegal, İspanya ve Arjantin’de,
K. pneumoniae dışındaki Enterobacteriaceae türlerinde de OXA-48 tanımlamıştır. Her ne
kadar Yunanistan’da KPC-2, NDM-1 ve VIM-2 karbapenemazlar baskın olsa da, 2012 yılında blaOXA-48 taşıyan K. pneumoniae kaynaklı bir salgın da bildirilmiştir (32).
2.3.2. Karbapenemaz üretiminin taranacağı fenotipler
Bakterilerde karbapenemazların klinik örneklerde saptanması, otomatik sistemler veya disk difüzyon duyarlılık test sonuçlarının dikkatli bir analizine dayanır. Otomatize sistemler karbapenemaz üreten tüm mikroroganizmaları güvenilir bir şekilde belirleyemeyebilir, tutarsızlıklar gözlenebilir. Ertapenem, karbapenemaz tespitinde duyarlılığı yüksek ancak özgüllüğü düşük bir antibiyotiktir. Meropenem, karbapenemazların saptanmasında duyarlılık ve özgüllüğü en iyi ajandır. EUCAST kılavuzuna göre, sınır değerler antibiyotik duyarlılığının sınıflandırılması için gereklidir. Ancak epidemiyolojik veri ve enfeksiyon kontrolü açısından karbapenemazların belirlenmesine yönelik özel testler önerilmektedir (102).
Rutin duyarlılık testlerinde karbapenem duyarlılığında azalma saptandıktan sonra karbapenemaz varlığı laboratuvarda fenotipik veya moleküler yöntemler ile tespit edilebilir. Fenotipik yöntemler arasında; inhibitör bazlı yöntemler (kombine disk testleri, kombine gradiyentli stripler), kolorimetrik yöntemler (CarbaNP testi, Blue-Carba testi ve -CARBA testi™ gibi), karbapenem inaktivasyon yöntemi (CIM), lateral akım yöntemi, kromojenik besiyerleri, Modifiye Hodge Testi ve MALDI-TOF MS yer alır (8, 103). Moleküler testler arasında; PCR, Klonlama ve sekanslama, Oligonükleotid hibridizasyonu, PGFE ve MLST yer alır (103).