A. O. Vet. Fak. Derg. 35 (2-3) : 218-226 1988
OMliRGASIZLARDA HORMONLAR
Baki Yılmaz
HOl'mone~ of im-crtebrates
Summar)': The development and meıamorphosis in inverlebrales is
(regulaled) by some hormones. These are prolllOraeicoıropic hormone
( PTTH), molling hormoııe (ecdysane), bursicon, juvenile hormone
( J H) and eclosion hormoııe. These are produced and secreled from cells in various parIs of ıhe organism. Proıhoracicoıropic hamıone (PTTH) sıimulales ıhe proıhoraeic glalld lo secrele ıhe molt-inducing faelor.
Proıhoracicoıropic hormone (brain hormone) acıing logeıher wiıh eedisone iniliales ıhe transformaıioıı of lana to pupa phase. Ecdisone, aetivates molting process and meıamorphosis in larval, pupal phases and iıı adulıhood. Bursieoıı, iııfluences certain aspecls of cll1icle develop-ment, including the process of tOlming (i,e hardening and darkening). Juvenile hormone, promotes the retenliaıı of the immature (ju venile) charaeteristics of the larva by pasIfJoning meıamorphosis until the adult slage.
It causes molıing in larvae a few ıimes iıı ıhe same phase and it also
makes ıhe larvae become bigger before it promotes the pupal phase.
Iı regulaıes molting, acıing logether with ~-ecdysone. In addition, ec-10sio/1hormone and bursicon have same functions at the last stage of /IIolting.
LLL ıhis article the formatian, seerelion and activaıion of the inver-tebrate hormones have been reviewed.
Özet: Omurgasızlarm gelişimi ve başkalaşımı birkaç hormon un
de-netimi altmdadır. Bunlar protorakotrop hormon (PTTH), deri
değiştir-me hormonu (ekdizon), bursikon hormonu, gençlik hormonu (GH) ve
eklozion hormonudur. Bunlar organizmal1ln çeşitli yerlerindeki hücre-lerde oluşur ve salmır.
OMURGASIZDA HORMONLAR 219
Protorakotrop hormon (PTTH) protoraks bezlerini uyanr ve deri
değiştirme hormonu salmmıına neden olur. Protorakotrop hormon
(be-yin hormonu) ekdizon hormonu ile birlikte larvadan pupa dönemine ge-çişi başlatır. Ekdizon, larva, pupa ve ergin dönemde deri değişimini ve başkalaşll111 sağlar. Bursikon kabuğlin hazı gelişme durumlanna,
sert-leşme ve renk koyulaşmasl11u etkir. Gençlik hormonu larvanm genç
k([lmaslJ11sağlar ve başkalaşll1l1 önler. Larvanl11 birkaç kez aym evrede
deri değiştirmesine ve pupa olmadan önce büyük boyutlara ulaşmasma
neden olur. Beta ekdizon ile hirlikte normal deri değişimini düzenler. Aynea eklozion ve hursikon hamıonu kabuk değişiminin son aşamasl11da görevalır.
Bu derlemede omurgasr::lartn hormonlannm oluşumu, salmımı ve
etkisi ana hatlanyla gözden geçirilmektedir.
Giriş
Omurgasız hayvanlarm bir çoğunda, omurgalılarda salman ep i-nefrin, asetilkolin ve kızgınlık hormonları ile karşılaştırılabilecek hor-monlar salındığı bilinmektedir (3, 6, 19, 24). Gerçekten bazı omurga-sızların doku özlerİ (ekstreieri) şırınga ile verildiğinde de epinefrin ve asetilkolinin etkisine benzer bir etki ortaya çıkmaktadır. Ayrıca, bir-hücrelilerin bazı türlerinde düşük derişimde (konsantrasyon) büyüme ve üremeyi hızlandıran, yüksek derişimde ise yavaşlatan düzenleyici hormonların bulunduğu belirlenmiştir (3, 5, iI, 15, 25).
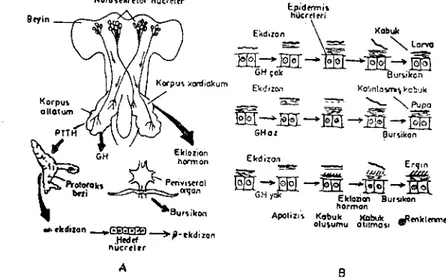
Böceklerin gelişimi ve başkalaşımı başlıca birkaç hormonun dene-timi altındadır (6, iI, i5). Bu hormonlar organizmanın çeşitli yer-lerinden salınır. Örneğin, nörosekretör hücreler omurgasızların beyin gangliyonunda bulunur (Şekil i - A). Hücregövdeleri pars interse-rebralis kesimindedir (6, 25). Bu hücreler protorakotrop hormon (PTTH) salar. Buna aynı zamanda beyin hormonu adı da verilir (3, 6, 15, 25). Protorakotrop hormon akzaplazma ile depolama yerlerine (nörohema) organlar) taşınır ve burada depo edilir. Son zamanlara kadar protorakotrop hormon depolayan organların korpus kardia-kum' lar olduğu sanılmaktaydı. Ancak, protorakotrop hormon salan nörosekretör beyin hücrelerinin korpus kardiakumlar'dan geçerek korpus allatum'da sona erdiği belirlenmiştir. Korpus allatum korpus kardiakum'un iki yanında bulunur. Böylece korpus allatum'daki nö-rosekretör hücre uçlarında oluşan PTTH, kana verilir (3, 6, 8, 9, 15,
Mo-220 B. YILMAZ
lekül ağırlığı yaklaşık 4500 dalton olan bir polipeptit olduğu bildiril-mektedir (6, 25). Nöro\tkrttö, hücreltr 8ryin
,-PTTH ~ GH Ekloı',on 'lı '5!.ıIı... hormon ~ Prolorok. ~-ı,~ Pm., ••,ol !>ni ~crqon "aunikonol-~kdiıon _ı:EIiE%L ~ o ••kd'
.tf~tf r' Ilon nucrtlrr A Ekdiıo~
-=
E,~,n ~ ,... 1.U4' £fa~-m-..m-E3-~
GHy~ EkioDerı 8u"lkon hormonApoli,,', Kabuk Kabule ..RMki...". olu~umu aıııması .'
B
Şekil 1. Böcek endokrin sistemi (A) ve kabuktaki değişimler (B) (6). Fig. 1. Insect endocrine system (A) and cuticular changes (B) (6).
Kana geçen PTTH protoraks bezini uyam ve başkalaşım ı sağ-layan alfa ekdizon'un salınımına neden olur (Şekil I-A, 2). Alfa ek-dizon kolesterol e ve omurgalılardaki steroit hormonların yapısına ben-zemektedir (3, 6, 8, i i, 14). Başlıca değişiklik, alfa ekdizon'da OH
HO
HO
Şekil 2. Q( - ekdizon (6).
Fig. 2 Q( - ccdysonc (6).
gruplarının daha çok bulunmasıdır. Bu gruplar, molekülü suda çö-zünür duruma getirir. Böceklerde, alfa ekdizon oluşabilmesi için besin-lerinde kolesterol bulunması gerekir. Alfa ekdizon, prohormon
ola-rak düşünülmektedir. Birçok çevresel hedef hücrelerde (yağ dokular) 20 - hidroksiekdizona dönüşür. Beyinden ya da ön gangliyondan gden uyarı protoraks bezinden ekdizon üretilmesine neden olur (6). Ekdizo-na deri değiştirme hormonu da denir. Larva, pupa ve ergin dönemde deri değişimini sağlar. Ekdizon hücrede adenilat siklaz-siklik AMP sistemine etkir. Bundan sonra çekirdeğe taşınan ve özel genleri etkin hale getiren reseptörlere bağlanarak epidermisi uyarır. Uyarı sonucun-da yeni kabuk oluşmaya başlar. Eski kabuk alttaki epidermis hüı;re-lerinden ayrılır (apolizis). Bunu yeni kabuk oluşumu izler (Şekil I-B). Yeni kabuk yumuşak olup, büyüme ve gelişmeye elverişlidir. Zamanla proteinlerle birleşerek sertleşir. Sertleşmeyi bursikon hormonu uyarıro Ayrıca, eski kabuk epidermisten salınan değişim sıvısında bulunan en-zimlerle de kısmen parçalanır (6, ll, 25). Larvalara çok az miktarda ekdizon hormonu şırınga ile verildiğinde i5 dakika içerisinde belirli bir kromozomun özel bir bölgesinin kabardığı belirlenmiştir (puf olu-şumu). Ekdizon gençlik hormonu ile birlikte normal deri değişimini düzenler (3, 6, iI, 15, 23, 25). Nitekim, gençlik hormonu ortamda yük-sek düzeyde bulunduğunda larva tipinde yeni kabuk gelişir. Gençlik hormonu düşük düzeyde ise, ergin tipinde kabuk oluşur ve ayrıca baş-kalaşım da gerçekleşir (Şekil I-B). Protorakotrop hormon ekdiıon ile birlikte pupa döneminde geçişi başlatır (6).
Beyinde ve sinir kordonundaki diğer nörosckretör hücrelerde bursikon hormonu oluşur. Bursikon molekül ağırlığı 2000~OOOO dalton olan bir polipeptittir. Adenilat siklazı etkin hale getirir. Kabu-ğun bazı gelişme durumlarını, sertleşmesini ve renk koyulaşmasını dü-zenler. Bu sertleşme ve renk koyulaşma olayı değişimden sonra birkaç saatte tamamlanır (6, 25).
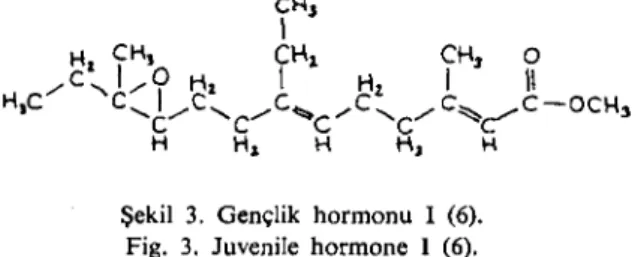
Beynin yanında yer alan ve sinirsel yapıda olmayan korpus alla-tum'lar gençlik hormonu (juvenil hormon) salar (Şekil 3). Ayrıca genç-lik hormonu (GH) bazı erkek güvelerin abdomenindeki yağ bezlerinde de üretilir (1-3, 5, 6). Bu hormon değişime uğramış, açık zincirli
ter-Şekil 3. Gençlik hormon u i (6). Fig. 3. Juvenile hormone ı(6).
222 B YILMAZ
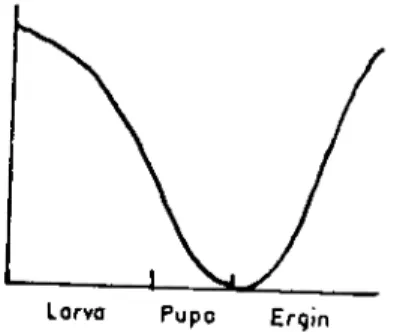
pen türevidir (25). Erken larva evresinde gençlik hormonu dolaşımda en yüksek düzeyde olup, son larva evresinde oldukça azalır ve pupa döneminin sonunda en düşük düzeye incr (6). Üreme yeteneği olan erginlerde gençlik hormonu salınımı yeniden artmaya başlar ve düzeyi yükselir (Şekil 4).
Pupa Erqin
Şekil 4. Böceklerde yaşam boyunca gençiik hormonu düzeyindeki degişimler (6).
Fig. 4. Changes in the levcl of ju\'cnile hormonc durinıı the insect life.
Anılan hormonun etki biçimi ekdizon ve diğer steroit hormon-larınki gibidir. En çok erken larva evresinde etkinlik gösteren gençlik hormonu başkalaşıma, diğer bir deyişle larvanın pLlpa ve ergin hale geçmesine engelolur (6, 7, 14, 16, 23). Fakat deri değişimini engelle-mez. Pupa dönemine gelmeden önce larvanın aynı evrede birkaç kez deri değişimini, büyük boyutlara ulaşmasını ve böylece larvanın genç kalmasını sağlar. Son larva evresinde gençlik hormonu salınımı aza-lır, ekdizon salınımı ise artar. Bu nedenle bu evrede ekdizon etki ba-kımından gençlik hormonuna baskın olmaya başlar (5). Gençlik hor-monu dolaşımdan kaybolunca da larvada başkalaşım oluşur. Böylece,
larva önce pupa, daha sonra da ergin hale gelir (5, 16, 17, 2 I, 22). Genç larvaların korpus aliatLlm'ları çıkarıldığında erken başka-laşım oluşur ve cüce olan ergin bir birey gelişir (5,9, 12). Buna karşın son evreye gelıniş larvaya fazla gençlik hormonu üreten genç larvanın korpus allatum'u aktarılırsa, larva ek deri değiştirmek üzere uyarılır ve pupa olmadan önce dev boyutlara ulaşır. Böylece dev yapılı bireyler oluşur ve gelişim normalden fazla sürer (5, 18, 20).
Üreme yeteneği olan ergin dişilerde gençlik hormonu düzeyi yeni-den yükseldiğinyeni-den üreme sisteminin gelişiminde de roloynar. Bu da yumurtanın olgunlaşmasını uyarır. Bazı böcek türlerinin erkeklerinde
erkeklik özelliklerinin gelişimine neden olur. Dişi böceklerde yurnur-talıklarda yumurta sarısı proteinlerinin oluşumunu da düzenler (4, 5, 6,
LO, 13, 25).
Eklozion hormonu korpus kardiakum'dan salınan ve molekül ağırlığı yaklaşık 8000 dalton olan bir polipeptittir (6, 25). Söz konusu hormon siklik guanilat siklazı etkiler ve siklik guanozin monofosfat (GMP) düzeyini önemli ölçüde yükseltir. Bursikon hormonu ile bir-likte kabuk değişiminin son evrelerini düzenler ve eski kabukların sıy-rılmasını başlatır (I i,24, 25). Yeni değişmiş kabuk yumuşak ve soluk renktedir. Bundan sonra bursikon etkisi altında kabuk sertleşir, renk koyulaş!r. Bazı türlerde pupa kabuklarının atılmasını eklozion hor-monu sağlar (6). Her kabuk değişiminde kabuğu n türü salınan gençlik hormonu düzeyine bağlıdır (6, 15, 25).
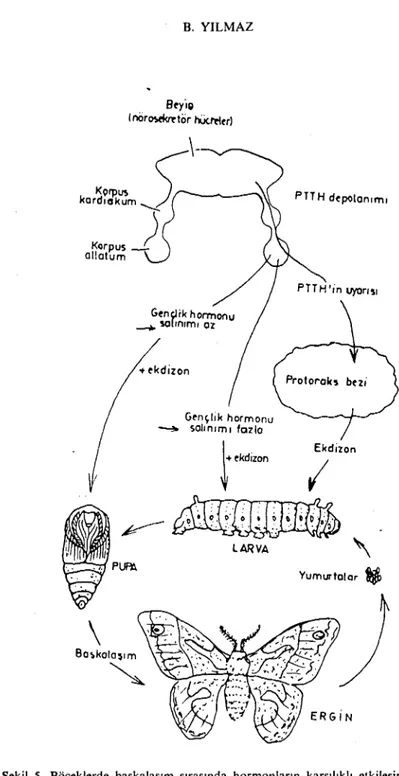
Şekil 5'de hormonların cecropia güvesinde büyüme ve gelişim üze-rine etkisi görülmektedir. PTTH, gençlik hormonu ve ~-ekdizon ı~vre değiştirmenin başlamasını etkiler. Larva değişmeleri protorakotrop hormon la (PTTH) uyarılır. PTTH protoraks bezinden ekdizon hor-monu salınımına neden olur (3. 6, 8, 25). Büyüme devam eder, ancak gençlik hormonu (GH) en düşük düzeyin altına düşmediği sürece bö-cek larva olarak kalır. Bu büyüme ve değişim 4 ya da 5 evrede olur. Bu sırada gençlik horınonunda sürekli bir azalım görülür. Gençlik hormonu düzeyinin azalımı pupa evresine geçişe olanak sağlar. Güve kışı pupa halinde geçirir. Sürekli soğuk etkisi altında kalma PTTH salınımıııı uyarır. Bu da ekdizon salınımına yol açar. Ekdizon, gençlik hormonu yokluğunda ergin güvenin oluşumuna neden olur. Bir bö-ceğin normal gelişimi her evrede gençlik hormonu düzeyi ile düzen-lenir. Gençlik hormonunun rolü, iki yaşayışlılar yani kurbağagiller-deki (amfibiler) tiroit hormonlarının sağladığı düzenlemeye benzer biçimdedir. Her iki durumda da, hormon düzeyi ile gelişme evresi ara-sında dengesizlik oluşursa, normalolmayan bir gelişim görülür (6, 7,
ii, 14, 17, 25).
Sonuç
Tüm omurgasızlarda hormonlar birbirinin taın benzeri değildir. Örneğin, I, Il, Illolarak üç tipi bulunan gençlik hormonunun L ve II. tipi daha çok kelebeklerde, IIi. tipi çekirgeler, kınkanatlılar ve zar-kanatlılarda bulunmaktadır. Protorakotrop hormon da A ve B olarak değişik türlerde rastlanılmaktadır. Ayrıca, literatürde böceklere
iliş-224 B. YILMAZ Bty'ı~ lnör~tör ~trl Yumurtalar
tııı
PH H dtpolanımı Gtnçıik hormonu salınımı fozlai.
Ekdizon W.tkdizon (~.~~fi
.•:.:.
lARVA " i P~ \-K,~~C--kordiCIkum ~
r
KorpusJ
ollatumŞekil 5. Böceklerde başkalaşım sırasında hormonların karşılıklı etkileşimi (6). Fig. 5. Hormone interactions in insects during the metamorphosis process (6).
kin hormon bildirimlerine çok rastlanılmasına karşın, diğer omur-gasızlarınkine ilişkin bildirimlere az rastlanılmaktadır. Sonuç olarak, omurgasızların, gelişim, başkalaşım konularında hormanlarla ilişki-sinin açıklanmasını bekleyen pek çok sorunun bulunduğunu belirte-biliriz.
Kaynaklar
i. Baker, F.C., Hagedorn, H.H., Schooley, D.A. and Wheelock, G. (1983). MG'sqııito jııvenile lıormoııe: Ideıııificatioıı and bioassay activity. J. Insect. Physiol., 29, 465-470.
Cited: Readio, J., Peck, K., Meola, R. and Dahm, K.H. (1988). Corpus allatum acıi-viıy (İlı vitro) iıı fen/Qle cıılex pipiens during adıılt life cycle .. J. Insect. Physiol.,34 (2): 131-135.
2. Bergot, B.J., Rateliff, M., Schooley, D.A.(ı98 I). Method for qııalltitative determiııatioıı of the four kılOlVIIjııveııile hormoııes iıı iıısect (İssııe usiııg gas c1lromatograpl:ymass spectrometry. J. Chromat. 204: 231-244. Cited: Readio, J., Peck, K., Meola, R. and Dahm, K.H. (1988). Corpus allarum activity (iııI"irro)iııfenıale culex pipieııs during adulr life cycle. J. Insect Physİol., 34 (2): 131-135.
3. Chen, J.H., Fugo, H., Nakajima, M., Nagasawa, H. and Suzuki, A. (1987). Neurohor-moııes in deretopiııg embryos of the silkll'orm, Bombyx mori: The presence aııd clıarac-teristin of prothoracicotropic Iwrmone. B.J. Tnsect. Physiol., 33 (6): 407-41
ı.
4. Demirsoy, A. (1985). Yaşamııı temel kuralları. Cilt I, kısım, i,Hacettepe Ünivı:rsitesiyayınları: A /52, Metaksan Ltd. Şti., Ankara.
5. Demirsov, A. (1985). Yaşamııı temel kl/ralla,.,. Cilt I, kısım, II. Hacettepe Üniversitesi yayınları: Ai 53, Metaksan Ltd. Şti., Ankara.
6. Eckert, R., Randall, D.(J983). Aııimal plıysiology. Second edition. W.H. Freeman and Company, Nev, York.
7. Feinsod, F.M. and Spielman, A.(J980). Indepeııdemly regulated juvenile hornıone acti-viry aııd ritellogcııcsis iıı mosquiroes. J. Insect. Physio1., 26: 829-832.
8. Fugo, H., Chen, .J.H., Nakaiima, M., Naga~awa, H. and Suzuki, A. (1987). Neurolzor-maııiııderelopillf{ cmbryos o( the silkworm. Boıııbyx ıııori: Prescııce and characteristics of PTTH-S. J. Insect. Physiol. 33: 243-248.
9. Gwadz, R.W. and Spjehnan. 1\.(1973). Corpus allatııııı control of orariaıı derelopment in Aedes aegypti. J. Insect. Physİol., 19: 1441-1448.
ıo. Hagedom, H.H., Tumer, S., Hagedom, E.A., Ponteeorvo, D., Greenbaum, P., Pfeiffer, D., Wheelock, G. and Flanagon, T.R. (1977). Postemergence grol;'tlı of the (IvOloian follicles of Aedes aegypti. J. Insect. Physiol., 23: 203-206.
1
ı.
Kcrkut, G.A., Gilbert, L.I. (1985). Comprelıensive bısect Physiology, BioclıemisrryalZd Plıarmac%gy. Vol. ı,7, 8. Pergamon Press, Oxford.12. ua, A.a. (1963). Same re/ariol/slıip.\' betll'eelZ elZviroıııııent, corpora allata, al/d egg matllratiolZ in acdil/e mosqııitoes. J. Insect. Physİol., 9: 793-809.
226 B. YILMAZ
13. Lea, A.O. (1968). Matiııg 'ritliout ilısemiııatiol1 inı'il'gin Aedes aegypti. J. Inseet. Phys.-ol., 14: 305-308.
14. Meola, R. and Readio, J. (J987) . .furel1ile lrormOlıe rcgulation of i!le sccoııd biling cycle
incıılex pipiens. J. Inscct.. Plıysiol., 33: 751-754.
15. NO\'ak, V.J.A. (1976). Insect lIOI'I/IOI1CS.Chapman and Hall, London.
16. Patterson, J.W. (1974). A comparisol1 0/ the morpllOgel1ctie al1d sıerili~ing activities of juvel1ife Irorll/one mill/ics 011Aedes aegypti. J. Insect. Physiol., 20: 2095-2106.
17. Readio, J. and Meola, R. (J985). T",o stages of jUl'eııile Irormo/l-mediated groırtır 0/ secol1dary lofliefes il1 cıdex pipiel1s. J. Insec1. Plıysiol., 3.1: 559-562.
18. Readio, J., Prck, K., Meola, R. and Dahnı, K.H. (1988). Corpııs aflatum aetivity (in
ı'itro) iıı/emale culex pipieııs dııring adult life cycle. J. Insect. Physiol., 34 (2): 13ı-135. 19. ROl'kstein, M. (1964). Tire p!lysiology o!'iııseera. Vol. I-III. Aeademic press. New York
and London.
2G Rossignol, P.A., Feinsod, F.M. and Spielman, A. (198ıL.Iıılribiıor)' rC'gulation of eorpus aflatum aetivity ;ıımosquiıoes. J. Inseet. Physiol.. 27: 651-654.
21. Ruegg, R.P., Tobe, S.S. and Loher, N. (1986). JIIl'e/lile lloymol1e /;ios)'lufıesis duril1g egg developmellt ilı the ericket, Teleogryflııs commodus. J. InsC<'t.Physiol., 32: 517-521. 22. Shayiro, A.B., Wheelock, G.D •• Ha~edorn, H.H., Baker, F.C., Tsai, L.W. and Schoo-ley. D.A. (ı986).Juvenile hOl'l/lOl1e al1d jıırel1ile lroımol1e esterase il1 adult females of tire mosquito Aedes aeg)pli. J. Inseet. Physiol., 32: 867-877.
23. Villee, C.A. (1979). Gel1el biyoloji. «ev. ~işli, M.N., Bozeuk, A.N., Bozcuk, S., Boş-gelmez, A.). Milli Eğitim Basımevi, IstanbuL.
24. Wigglesworth, V.B. (1974). Tire pril1ciples oj il1seet plrysiology. 7. cd., Methwen, Lon-don.