Contents lists available atScienceDirect
Industrial Crops & Products
journal homepage:www.elsevier.com/locate/indcropMultidirectional insights on Chrysophyllum perpulchrum leaves and stem bark
extracts: HPLC-ESI-MS
n
pro
files, antioxidant, enzyme inhibitory,
antimicrobial and cytotoxic properties
Mehmet Cengiz Baloglu
a,⁎, Eulogio J. Llorent-Martínez
b, Muhammad Zakariyyah Aumeeruddy
c,
Mohamad Fawzi Mahomoodally
c, Yasemin Celik Altunoglu
a, Buket Ustaoglu
a, Mustafa Ocal
a,
Songül Gürel
d, Kouadio Bene
e, Kouadio Ibrahime Sinan
f, Gokhan Zengin
f,⁎aDepartment of Genetics and Bioengineering, Faculty of Engineering and Architecture, Kastamonu University, Kastamonu, Turkey bDepartment of Physical and Analytical Chemistry, University of Jaén, Campus Las Lagunillas S/N, E-23071, Jaén, Spain cDepartment of Health Sciences, Faculty of Science, University of Mauritius, 230, Réduit, Mauritius
dDepartment of Biology, Faculty of Art and Science, University of Bolu Abant Izzet Baysal, Bolu, Turkey
eLaboratoire de Botanique et Phytothérapie, Unité de Formation et de Recherche Sciences de la Nature, 02 BP 801 Abidjan 02, Université Nangui Abrogoua, Abidjan, Cote
d’Ivoire
fDepartment of Biology, Science Faculty, Selcuk Universtiy, Campus, Konya, Turkey
A R T I C L E I N F O Keywords: Africa Antioxidant Enzyme inhibition Polyphenolic Antimicrobial Anticancer A B S T R A C T
Chrysophyllum perpulchrum Mildbr. ex Hutch. & Dalziel has been reported to possess several therapeutic prop-erties in African traditional medicine. However, its pharmacological propprop-erties have not been fully studied. Herein, we focused on the evaluation of the antioxidant, enzyme inhibitory, antimicrobial and cytotoxic effects of three solvent extracts (ethyl acetate, methanol, and water) of the stem bark and leaves. Polyphenolic com-ponents of the extracts were also identified by high-performance liquid chromatography with electrospray io-nization mass spectrometric detection (HPLC-ESI-MSn). The methanolic stem bark extract possess the highest
DPPH (2,2-diphenyl-1-picrylhydrazyl) scavenging, ferric and molybdenum (VI) reducing, and the highest in-hibitory effect against acetylcholinesterase and butyrylcholinesterase. The aqueous stem bark extract displayed the highest ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid)) scavenging and cupric reducing power. In contrast, the leaf ethyl acetate extract was the most effective metal chelator and α-amylase inhibitor while the leaf methanolic extract exhibited the highest tyrosinase andα-glucosidase inhibition. The highest total phenolic content (131.58 mg GAE/g extract) was recorded from the methanolic stem bark extract while the leaf methanolic extract was the richest in totalflavonoid content (30.43 mg RE/g extract). Flavan-3-ols were main group in the methanol extracts. The minimum inhibitiory concentration values for the methanolic stem bark extract against Gram positive and negative bacteria strains ranged between 62.5–125 μg/ml. Bark extracts of C. perpulchrum were very effective against Salmonella kentucky, Proteus vulgaris and Staphylococcus aureus at 62.5μg/ml. In addition, cytotoxic activity of stem bark samples was evident at lower concentration than those of leaf extracts. Stem bark extracts displayed the highest prevention against to HeLa cell line (IC50 =264μg/mL
after 48 h). In light of the above, C. perpulchrum might provide health benefits against oxidative stress and the associated chronic diseases. Nonetheless, the detailed mechanism of action is yet to be further investigated.
1. Introduction
Antioxidative compounds can manage oxidation process and thus natural dietary antioxidants have great importance tofight oxidative stress, which is known as imbalance between antioxidant and free ra-dicals. Plants or plant products have been known as potential sources of
antioxidants as witnessed in the literature. In this line, the search of new antioxidants from plants have been become one of the most at-tractivefield of research among the scientific community (Kasote et al., 2015).
Enzyme inhibition is another major strategy for the design of novel drugs. Though enzymes are essential for many biochemical processes in
https://doi.org/10.1016/j.indcrop.2019.03.066
Received 4 February 2019; Received in revised form 26 February 2019; Accepted 26 March 2019
⁎Corresponding authors.
E-mail addresses:[email protected](M.C. Baloglu),[email protected](G. Zengin).
Available online 01 April 2019
0926-6690/ © 2019 Elsevier B.V. All rights reserved.
the body system, abnormally high enzyme activity can lead to major pathologies. Consequently, the modulation of such enzymes via natural inhibitors has been crucial for the prevention and treatment of hy-pertension, control of inflammatory response, intervention in cell growth and cell cycle, and other disorders (Future Market Insights, 2018). More than 90 enzyme inhibitors are present in the market as drugs and a greater number is under development and drug develop-ment pipeline. By 2022, the global market for enzyme inhibitors is expected to reach $179.9 billion from $168 billion in 2017, at a com-pound annual growth rate of 1.4%, from 2017 to 2022 (Dewan, 2018). Plants are excellent sources of natural antioxidants and possess a wide array of metabolites with significant enzymatic properties. Furthermore, metabolites isolated from plants represent an epitome for human remedies and on many occasions have proved to be good can-didates for promising drug development against a panoply of diseases. The African continent is immensely rich in biodiversity filled with 40,000–45,000 plant species, among which 5000 are used as traditional medicines (Mahomoodally, 2013). The genus Chrysophyllum from the family Sapotaceae is a major group in Angiosperms consisting of 89 accepted species names, 251 synonyms and 18 unassessed summing up to 358 scientific species names (The Plant List, 2013). Chrysophyllum perpulchrum Mildbr. ex Hutch. & Dalziel is a tall tree reaching a height of 40 m with a huge, rectilinear and cylindrical trunk, 100 cm in dia-meter. The base of the tree is distinctivelyfluted and buttressed (Bidie et al., 2016; Tropical Plants Database, 2018). It is mainly found in tropical countries of Africa including Liberia, Cote D'Ivoire, Ghana, Nigeria, Cameroon, Gabon, Central African Republic, Congo, Uganda, and Tanzania. This species prefers habitat such as semi-deciduous forest, most abundantly in dry forest types, particularly on slopes. In Eastern Africa, it is present in rainforests at elevations of 800-1,200 m (Tropical Plants Database, 2018). The wood of C. perpulchrum can be used for construction, flooring, furniture, musical instruments, car-vings, veneer and playwood. The latex obtained from the bark is used as a rubber substitute (Tropical Plants Database, 2018).
Chrysophyllum perpulchrum has several therapeutic properties in traditional medicine. Powdered roots are applied to swelling, and de-coction of the root is used to treat dysmenorrhea, female sterility, and schistosomiasis. Bark decoction prepared from the trunk is used to treat gonorrhea, abdominal pain, and skin conditions, while the leaves are used in the treatment of snake bites (Bidie et al., 2016). Moreover, decoction of the stem bark of C. perpulchrum is taken orally to treat malaria by the Bete populations of Issia (Central West Forest of Côte-d’Ivoire) (Guédé et al., 2010). In Tanzania, the roots are used for the treatment offlu (Choi et al., 2015).
With regards to its pharmacological properties, C. perpulchrum ex-tract was found to display quenching of DPPH and inhibition of lipid peroxidation. Catechin and two dimeric procyanidins were reported as the main compounds and the observed abilities can be attributed to these compounds (Philippe et al., 2010). The ethanolic extract also showed in vitro antiplasmodial activity against Plasmodium falciparum (IC50= 12.8μg/ml) (Guédé et al., 2010). The methanolic extract of C. perpulchrum bark also normalized blood glucose, insulin secretion, and lipid profile in diabetic rats (Philippe et al., 2016). Also, in a previous in vivo study (Bidie et al., 2016), treatment of rats with the aqueous ex-tract of C. perpulchrum (at doses 2000 and 5000 mg/kg bw) increased serum levels of blood platelets, white blood cells, high-density lipo-protein, and decreases low-density lipoprotein indicating its im-munostimulatory and cardioprotectory activity. In addition, pre-liminary phytochemical screening also revealed the presence of sterols, polyterpenes, polyphenols, tannins,flavonoids, alkaloids, and saponins in this species (Bidié et al., 2011).
With the aim to further explain the bio-pharmaceutical potential of this species, we focus on the antioxidant, enzyme inhibitory, and an-timicrobial-cytotoxic effects of several solvent extracts (ethyl acetate, methanol, and water) of the leaf and stem bark of C. perpulchrum, and also identified their polyphenolic components through HPLC-ESI-MSn
analysis. To the best of our knowledge, for thefirst time, in-depth in vitro biological properties of this species are explored using an array of biological assays.
2. Materials and methods
2.1. Plant materials and extraction procedure
Chrysophyllum perpulchrum was collected in Haut sassandre of Ivory Coast in 2018 and authenticated by the botanist Dr. Bene Kouadio in the Université Nangui Abrogoua, Ivory Coast. Stem barks and leaves were allowed to shade dry at room temperature for 10 days and grinded intofine powders.
Crude ethyl acetate and methanol extracts were prepared by ex-haustively extracting 5 g of ground plant material into 100 ml solvents overnight at room temperature. The extracts werefiltered and con-centrated in vacuo in a rotary evaporator at 40 °C. Crude water extracts were prepared using infusion method by boiling of 5 g plant material in 100 ml distilled water for 20 min. Thefiltrates were dried in a lyo-philizer. The residues were kept at + 4 °C until further analysis. 2.2. Profile of total bioactive compounds
With reference to our previous studies (Lazarova et al., 2014), the total phenolic and flavonoid content (by standard Folin-Ciocalteu method) and flavonoids (TFC) (by AlCl3 method) were determined; (gallic acid (mg GAE/g) for total phenolic content and rutin (mg RE/g) for totalflavonoid content, respectively were used to express the re-sults.
2.3. Chromatographic conditions
Chromatographic separation was performed by using Agilent Series 1100 (combined with G1315B diode array detector). The separation was done with a polar colunm (Luna Omega Polar C18 analytical column of 150 x 3.0 mm and 5μm particle size). The chromatographic system was connected to an ion trap mass spectrometer (Esquire 6000, Bruker Daltonics) equipped with an electrospray interface operating in negative ion mode. The HPLC-MS conditions were previously reported (Llorent-Martinez et al., 2018). 5 mg of dried extract (DE) was re-dis-solved in 1 ml MeOH,filtered through 0.45 μm PTFE membrane filters, and 10μL was injected into HPLC.
2.4. Evaluation of biological activities
Tyrosinase, α-amylase, α-glucosidase, and cholinesterases were identified as target enzymes and the protocols for these bioassays were described in our previous publications (Lazarova et al., 2015). Standard inhibitors (acarbose (against α-amylase and α-glucosidase), galanta-mine (against or AChE and BChE), and kojic acid (against tyrosinase)) were used to express the enzyme inhibitor properties.
Antioxidant capacity of the extracts were spectrophotometrically screened by different experiments as ferrozine assay (for chelating abilities), Phosphomolybdenum, reduction potentials (by FRAP and CUPRAC assays) and radical attenuation (using DPPH and ABTS radi-cals). Standard compounds (TE/g and EDTAE/g) were used to express the antioxidant properties and the detailed protocols are describe in the literature (Lazarova et al., 2014).
2.5. Statistical evaluation for total bioactive components, antioxidant and enzyme inhibitory properties
To observe differences among extracts, One-way ANOVA was se-lected with Tukey’s test (p < 0.05). All procedures were performed by GraphPad Prism 7 software (GraphPad Software, San Diego, California, USA).
2.6. Determination of MIC and MBC
Twelve bacteria (six Gram positive and negative strains) were used to detect antimicrobial evaluation of C. perpulchrum bark and leaves extracts. These bacterial strains were provided by Department of Genetics and Bioengineering of Kastamonu University (Kastamonu, Turkey). Minimum inhibitory concentrations were defined as the minimum concentrations that prevent bacterial growth under certain growth conditions (Andrews, 2002). At the beginning, bacterial broth cultures were prepared to have 0.5 McFarland density. Three plates were used for leaf and bark extracts of the plant. 100μl growth medium was added to each well in a 96-well plate. Then, two-fold serial dilu-tions were performed from plant extracts at range of 15.6/500μg concentration. 100μl solution was discarded from last dilution. 10 μl from 0.5 McFarland bacterial broth cultures was added to the wells. Positive control included bacteria and growth medium while negative control was included only growth medium. The plates were incubated at 37 °C overnight. After incubation, turbidity was determined using a spectrophotometer at 600 nm wavelength. MIC is the lowest con-centration where no growth is visually observed. The MBC test detect the lowest antimicrobial compound concentration which kills micro-organisms. To determine the MBC, the dilution representing the MIC and two different concentrations were plated. The plates were again incubated at 37 °C overnight. MIC and MBC values were determined according to these measurements.
2.7. Cytotoxic assessment 2.7.1. Cell culture maintenance
HeLa cancer cell line was supplied by Boğaziçi University, Department of Molecular Biology and Genetics (Istanbul, Turkey). Culture media of these cell lines were prepared in DMEM solution (1% penicillin-streptomycin (Pen-Strep) and 10% fetal bovine serum (FBS) and 1% non-essential amino acid). This medium was kept at + 4 °C. Cancer cell lines were maintained in an incubator (5% carbon dioxide and 37 °C). After the cells reached confluence, the cells were first wa-shed with 1X PBS and treated with 0.25% trypsin-EDTA solution and then cells were scraped with cell scraper. The cells were collected by centrifugation. After cells were resuspended in culture medium, they were seeded into three 96-well plates.
2.7.2. MTT assay and calculation of IC50value
HeLa cells (10*4) were seeded in 96-well plates and grown for 24 h. Cells were treated with variable doses of C. perpulchrum extracts (62.5, 125, 250, 500, 1000μg/ml in DMSO/water (1:5)) for 48 h. Then, the medium was renewed with DMEM (with 0.5% FBS + 0.5 mg/ml MTT) and kept at 37 °C for 4 h. Cells were mixed with as solution containing 40 mM HCl/isopropanol+ 3% SDS for 15 min and the MTT-formazan crystals were dissolved by this solution. The absorbance of samples was measured using microplate reader at 570 nm (Multiskan Go, Thermo Scientific, USA). Half maximal inhibitory concentration known as IC50 was determined using the log (inhibitor) versus normalized re-sponse–variable slope analysis function of GraphPad Prism 7 software (GraphPad Software, San Diego, California, USA).
2.8. Multivariate analysis
Pearson correlation and hierarchical cluster (heat map) were per-formed to detect possible relation between bioactive compounds (phenolics andflavonoids) and biological properties (antioxidant and enzyme inhibitory effects). Also, Sparse Partial Least Squares (sPLS-DA) and principal component (PCA) analysis were selected to provide a further insights on the extracts. All procedures were performed by R software v. 3.5.1.
3. Results and discussion
3.1. Total amounts of phenolics andflavonoids
The C. perpulchrum leaf and stem bark extracts were determined with regard to TPC and TFC and thefindings are presented inFig. 1. In the stem bark extracts, the methanol extract (131.58 mg GAE/g) was found to be the highest TPC, followed by the aqueous (68.74 mg GAE/ g) and ethyl acetate extract (36.63 mg GAE/g). Regarding the leaf ex-tracts, the methanol one also had the highest TPC (95.42 mg GAE/g) while the minimun level was expressed in the ethyl acetate one (41.27 mg GAE/g).
With regards to TFC, the methanolic leaf extract showed the best level (30.43 mg RE/g) when compared with other extracts (6.40 and 5.76 mg RE/g for the ethyl acetate and aqueous extract, respectively). Similarly, among the stem bark extracts, the methanol one possessed the highest TFC (2.42 mg RE/g) while the lowest amount was observed in the aqueous extract.
3.2. HPLC-ESI-MSn
The characterization of the phytochemicals in the methanol extracts (selected based on its overall higher biological activity compared to the other solvent extracts) was carried out by HPLC-ESI-MSnwith the ne-gative ion mode. The characterization of the compounds was performed with analytical standards as well as related scientific literatures. The characterization is reported inTable 1. Thesefindings are the first for C. perpulchrum leaves and stem barks. Following is a discussion of the results obtained.
3.2.1. Flavonoids
Flavan-3-ols were detected as mainflavonoids, which is consistent with the scarce studies performed in Chrysophyllum species. For in-stance, (epi)catechin, gallocatechin, and (epi)gallocatechin compounds
Fig. 1. Total bioactive components of the tested extracts. (Values expressed are means ± S.D. of three parallel measurements. GAE: Gallic acid equivalent; RE: Rutin equivalent; EA: Ethyl acetate; MeOH: Methanol. Different letters indicate differences in the extracts (p < 0.05).
have been reported in C. cainito L. (Amelin et al., 2018).
Epicatechin (compound11) was confimed with an analytical stan-dard. Compound5, with [M−H]−at m/z 609 and MSnbase peaks at m/z 441 and 423, was characterized as a prodelphinidin dimer B-type formed by two units of (epi)gallocatechin (Bresciani et al., 2015). Compound6 was also a prodelphinidin B, but consisting of (epi)ca-techin and (epi)galloca(epi)ca-techin (Friedrich et al., 2000). Compound 7 presented fragment ions characteristic of (epi)catechin at m/z 289, 245, and 205; hence, it was tentatively characterized as a derivative. Com-pounds9 and 22 were (epi)catechin dimers, whereas compound 13 was a trimer; all of them displayed neutral losses of 288 units, indicative of (epi)catechin (Kajdžanoska et al., 2010). Compound12, with [M−H]− at m/z 457, displayed fragment ions at m/z 169 and 125 (gallic acid) and was characterized as (epi)gallocatechin-O-gallate (Bresciani et al., 2015). Compound 14, with deprotonated molecular ion at m/z 561, showed a fragmentation pattern accord with (epi)afzelechin-(epi)
catechin (Kajdžanoska et al., 2010). Compound21 suffered the neutral loss of 152 Da (441→289), which corresponded to a galloyl moiety; hence, it was characterized as (epi)catechin-O-gallate.
Compounds 10 and 15, with deprotonated molecular ion at m/z 419, suffered the neutral loss of 162 Da (hexoside), and additional losses of 15 Da (methyl), so they were tentatively characterized as methylatedflavonoid-O-hexoside isomers.
Compounds16, 18, and 23 exhibited the deprotonated molecular ion at m/z 435, and MSnfragment ions at m/z 309, 303, 285, 151, and 125. The fragment ion at m/z 303 corresponded to taxifolin (after the loss of a pentoside moiety). This is accordance with taxifolin-O-pento-side (Hvattum, 2002;Mämmelä, 2001), which has not been previously reported in Chrysophyllum species.
Compound20 presented [M−H]−at m/z 461 and suffered the loss of 144 Da to yield myricetin at m/z 317, so it was tentatively char-acterized as a derivative.
Table 1
Characterization of the compounds found in the methanol extracts of C. perpulchrum.
No. tR(min) [M-H]−m/z m/z (% base peak) Assigned identification Leaves Stem bark
1 1.7 377 MS2[377]: 341 (100), 179 (20) MS3[377→341]: 179 (100), 161 (43), 143 (40), 113 (12) Disaccharide (HCl adduct) ✓ ✓ 2 1.7 317 MS2[317]: 225 (100), 207 (42), 165 (80), 125 (19) MS3[317→225]: 165 (84), 135 (100) Unknown ✓ 3 2.0 191 MS2[191]: 173 (85), 111 (100) Citric acid ✓ 4 3.1 358 MS2[358]: 179 (59), 161 (10), 143 (6), 131 (100), 119 (13) Hexoside derivative ✓ 5 3.5 609 MS2[609]: 591 (5), 483 (10), 441 (100), 423 (75), 305 (18) MS3[609→441]: 423 (100), 273 (7)
Prodelphinidin dimer B-type [2 units of (epi)GC] ✓
6 5.6 593 MS2[593]: 425 (100), 407 (67), 289 (28) Prodelphinidin dimer B-type [(epi)GC-(epi)
catechin] ✓ 7 6.4 451 MS2[451]: 405 (100), 289 (87) MS3[451→405]: 245 (93), 205 (100), 151 (89) (Epi)catechin derivative ✓ 8 7.1 321 MS2[321]: 225 (63), 183 (100) Unknown ✓ ✓
9 10.7 577 MS2[577]: 451 (15), 425 (100), 407 (53), 289 (37), 287 (24) (Epi)catechin dimer (B-type) ✓
10 11.3 581 MS2[581]: 419 (100) MS3[581→419]: 404 (100), 389 (14), 373 (45) Methylatedflavonoid-O-hexoside ✓ 11 12.1 289 MS2[289]: 245 (100), 205 (54), 203 (20), 179 (19) Epicatechin ✓ ✓ 12 13.2 457 MS2[457]: 331 (21), 319 (8), 305 (10), 193 (12), 169 (100) MS3[457→169]: 125 (100) (Epi)gallocatechin-O-gallate ✓ 13 14.1 865 MS2[865]: 739 (27), 695 (100), 577 (41), 425 (32), 407 (50), 289 (19), 287 (37)
(Epi)catechin trimer (B-type) ✓
14 14.8 561 MS2[561]: 435 (27), 425 (22), 407 (15), 289 (100), 271 (18) MS3[561→289]: 245 (100), 205 (21), 203 (21), 179 (34) (Epi)afzelechin-(epi)catechin (B-type) ✓ 15 15.5 581 MS2[581]: 419 (100) MS3[581→419]: 404 (100), 389 (10), 373 (41) Methylatedflavonoid-O-hexoside ✓ 16 17.1 435 MS2[435]: 309 (28), 303 (97), 285 (100), 151 (88), 125 (4) MS3[435→303]: 285 (100), 125 (17) Taxifolin-O-pentoside ✓ 17 18.0 471 MS2[471]: 169 (100) MS3[471→169]: 125 (100)
Gallic acid derivative ✓
18 19.0 435 MS2[435]: 309 (16), 303 (73), 285 (100), 151 (39), 125 (6) Taxifolin-O-pentoside ✓ 19 19.3 497 MS2[497]: 461 (100) MS3[497→461]: 229 (26), 199 (100), 169 (45) Unknown ✓ 20 19.7 461 MS2[461]: 317 (100) MS3[461→317]: 271 (100), 179 (37), 151 (16) Myricetin derivative ✓ 21 19.7 441 MS2[441]: 289 (100), 193 (11), 169 (27) MS3[441→289]: 245 (100), 205 (15), 203 (17) (Epi)catechin-O-gallate ✓
22 20.1 577 MS2[577]: 575 (215), 451 (26), 425 (100), 407 (66), 289 (20) (Epi)catechin dimer (B-type) ✓
23 21.6 435 MS2[435]: 309 (13), 303 (56), 285 (100), 151 (41) MS3[435→303]: 285 (100), 125 (22) Taxifolin-O-pentoside ✓ 24 22.2 433 MS2[433]: 301 (100) MS3[433→301]: 179 (64), 151 (100) Quercetin-O-pentoside ✓ 25 23.8 433 MS2[433]: 301 (100) MS3[433→301]: 179 (67), 151 (100) Quercetin-O-pentoside ✓ 26 24.5 447 MS2[447]: 301 (100), 151 (10) MS3[447→301]: 179 (100), 151 (67) Quercetin-O-deoxyhexoside ✓ ✓ 27 25.9 419 MS2[419]: 287 (100), 269 (54), 259 (36), 179 (11) MS3[419→287]: 259 (100) Dihydrokaempferol-O-pentoside ✓ 28 30.2 843 MS2[843]: 797 (100) MS3[843→797]: 503 (100), 485 (10) MS4[843→797→503]: 485 (100), 443 (25), 439 (17) Unknown ✓ 29 33.9 501 MS2[501]: 483 (31), 453 (100) Unknown ✓ ✓ 30 35.6 301 MS2[301]: 179 (97), 151 (100) Quercetin ✓ (epi)GC: (epi)gallocatechin.
Compound27, [M−H]−at m/z 419, suffered the neutral loss of 132 Da (pentoside) to yield the aglycone at m/z 287, which corre-sponded to dihydrokaempferol due to the fragment ion at m/z 259 (Fischer et al., 2011). Hence, it was characterized as dihy-drokaempferol-O-pentoside.
Quercetin (compound 30) was identified by comparison with an analytical standard. It was only detected in leaves extracts. Compounds 24 and 25, only detected in leaves, corresponded to quercetin-O-pen-toside isomers, due to the loss of 132 Da (penquercetin-O-pen-toside) to yield quercetin at m/z 301. Compound 26 was characterized as quercetin-O-deox-yhexoside due to the loss of 146 Da, and it was present in both leaves and stem bark. Both quercetin and its deoxyhexoside glycoside have been previously reported in C. canito fruits (Amelin et al., 2018), al-though pentoside glycosides are here mentioned in this species for the first time.
3.2.2. Other compounds
Compound1 corresponded to a HCl adduct of a disaccharide due to the [M−H]−at m/z 341 and fragment ions at m/z 179, 161, 143, and 113 (Gómez‐Caravaca et al., 2016). Compound4, with similar frag-mentation pattern, was tentatively characterized as a hexoside deriva-tive.
Compound3 was confirmed as citric acid due to the deprotonated molecular ion at m/z 191 and fragment ions at m/z 173 and 111 (Mena et al., 2012).
Compound17 was tentatively characterized as a gallic acid deri-vative according to the 169→125 fragmentation, typical of gallic acid. 3.3. Multivariate analysis
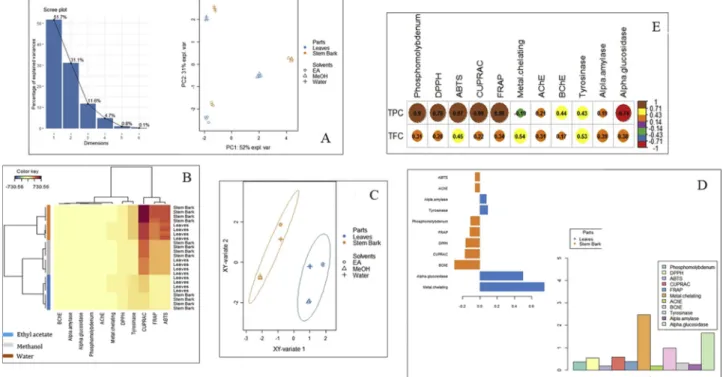
A preliminary unsupervised method such as principal component and hierarchical cluster analysis together with supervised sPLS-DA (Sparse Partial Least Squares Discriminant Analysis) were performed on the biological activities data to allow a first exploration of the major sources of variation in the data. These multivariate statistics are helpful approaches for a better visualization and understanding of highly di-mensional data sets. Both unsupervised principal component and hierarchical cluster analysis reveal differences between samples ac-cording to the type of solvent used for the extraction (Fig. 2A). Thefirst group included the ethyl acetate extracts of both studied parts, the second was represented by methanol extracts while the third group consisted in the water extracts. Thesefindings pointed out that what-ever the part of C. perpulchrum, the biological activities were affected by the extraction solvent used. The eigenvalue plot of PCA indicated that three components are sufficient to explain most information from the data (Fig. 2A). In addition, all replications of the same extract (part*-solvent) clustered together on heatmap, hence experimental conditions adopted were suitable (Fig. 2B).
On the contrary of unsupervised methods that we applied in order to uncovered differences between clusters without supervision, the su-pervised modelling sPLS-DA was performed considering the different studied parts as class membership criteria. In fact, PCA and HCA are unsupervised analyses where no information about the studied parts was provided in the method. Visualizing the sPLS-DA individual-plot, it was clear that the class prediction model discriminated the different extracts of C. perpulchrum into two distinct groups based on parts (Fig. 2C). This result indicates that the leaves and stem bark had some different profile depending on biological activities. In fact, the VIP analysis showed that the small subset of biological activities that better explain the differences across the leaves and stem bark extracts was metal chelating activity and alpha-glucosidase inhibitory (Fig. 2D). The performance of the sPLS-DA model was excellent, with R2X and R2Y values of 0.99 and 09.8, while Q2Y was 098. The sPLS-DA pattern clearly differentiated between the two parts (leaves and stem bark), while the unsupervised PCA and HCA methods suggested that among each parts there were some differences in terms of solvents.
3.4. Antioxidant activity
In the current study, we investigated the antioxidant capacities of C. perpulchrum leaves and stem bark extracts by assaying their DPPH and ABTS scavenging capacity, ferric and cupric reduction activity, metal chelating activity, and phosphomolybdenum assay (total antioxidant capacity) (Table 2).
By scavenging free radicals, antioxidants can enhance the stability and quality of food products and drugs, and terminate peroxidation chain reactions (Huyut et al., 2017). Among the different solvent ex-tracts of the stem bark of C. perpulchrum tested in this study, the me-thanol extract was found to be the best DPPH scavenger (73.23 mg TE/ g). Similarly, among the extracts of the leaf, the methanol extract showed the best activity (72.98 mg TE/g). The ethyl acetate extracts (for stem barks and leaves) displayed the weakest activity. As for the ABTS assay, the aqueous extract obtained from the stem bark exhibited the highest ABTS scavenging effect (490.71 mg TE/g) followed by the leaf aqueous extract (440.32 mg TE/g). The tested ethyl acetate extracts possessed the lowest ABTS quenching abilities.
Reducing power is considered as one of the most important anti-oxidant mechanisms and this is reflected to the electron-donating ca-pacity of antioxidant compounds. In this study, the reducing power of the plant extracts was determined by the FRAP and CUPRAC methods. The FRAP assay is based on the reduction of ferric to ferrous. An in-crease in absorbance indicates stronger reducing power of the extracts (Turan et al., 2016). In the current study, we found that the methanol extract of the stem bark of C. perpulchrum, followed by that of the leaf, displayed the highest ferric reducing power (440.30 and 352.96 mg TE/ g, respectively). The tested ethyl acetate extracts displayed the lowest activity. On the other hand, the CUPRAC method is based on the re-duction of Cu2+to Cu+at pH 7 in aqueous ethanol in the presence of neocuproine (2,9-dimethyl-1,10-phenanthrene) (Huyut et al., 2017). In the stem bark extracts, the methanol extract showed the strongest cupric reducing power (717.16 mg TE/g). As for the leaf, the ethyl acetate extract exhibited the lowest activity in the CUPRAC (172.86 mg TE/g). In general, the leaves and bark methanol extracts showed best activity for all antioxidant assays (Table 2).
Unlike the CUPRAC and FRAP methods, it forms a green phospho-molybdenum complex without any induction of free metal ions solution in the phosphomolybdenum assay. As shown inTable 2, it was observed that the methanolic extracts of C. perpulchrum exhibited the highest Mo (VI) reducing effect with the stem bark extract (4.00 mmol TE/g ex-tract) being more effective than the leaf (3.35 mmol TE/g).
Furthermore, we also assess the metal chelating effect of the solvent extracts of C. perpulchrum. Transition metal ions can be involved the production of free radicals by Fenton reaction. In this sense, the che-lating of these metal ions by antioxidant compounds is an important way in antioxidant mechanism (Phatak and Hendre, 2014). As can be seen inTable 2, among the leaf extract of C. perpulchrum, the ethyl acetate extract was the most effective metal chelator (45.49 mg EDTAE/ g). Regarding the stem bark extracts, the aqueous extract had the best metal chelating activity (10.94 mg EDTAE/g), while the ethyl acetate one did not display any effect in this assay.
From the abovefindings, we found that the considerable activity of the stem bark and leaf methanolic extract in the DPPH, FRAP, and phosphomolybdenum assay was in conformity with their levels of TPC and TFC since the highest amount was detected in methanol compared to the counterpart solvent extracts (ethyl acetate and water) (Fig. 2E). Indeed, several researchers have been reported that the positive cor-relation of TPC and TFC of plant extracts and their DPPH scavenging effect (Deng et al., 2015;Yang et al., 2015), ferric reducing (Yang et al., 2015; Zhao et al., 2015), and molybdenum (VI) reducing power (Arumugam et al., 2018; Wan et al., 2011). It is noted that several compounds identified in the methanolic extract by HPLC-ESI-MSn analysis, including (epi)catechin, quercetin, taxifolin, prodelphinidin, and (epi)gallocatechin, are known to possess antioxidant properties as
proven by several studies (Ozgen et al., 2016; Plumb et al., 2002; Shanmugam et al., 2017;Topal et al., 2016).
3.5. Enzyme inhibitory activity
In the present study, two cholinesterases (AChE and BChE), tyr-osinase, α-amylase, and α-glucosidase were selected to evaluate the enzyme inhibitory properties of C. perpulchrum extracts and the results are shown inTable 3.
Cholinesterase inhibitors are known to improve the cognitive function by inhibiting the enzymes that degrade acetylcholine and this approach is known as cholinergic hypothesis. Cholinesterase inhibitors have always been the standard approach to the symptomatic treatment of Alzheimer's disease (Grossberg, 2003). As shown in Table 3, the methanol extract of both parts of C. perpulchrum were the most effective AChE inhibitor (5.21 and 4.90 mg GALAE/g for the stem bark and leaf, respectively). Similarly, the methanol extracts showed the highest BChE inhibition (0.94 and 0.69 mg GALAE/g for the stem bark and leaf, re-spectively). In contrast, the weakest AChE and BChE inhibitors were the aqueous extracts.
New therapeutic approaches are also being explored to regulate postprandrial glucose levels because of the severe side-effects linked to commercially available anti-diabetic medications (Llorent-Martínez et al., 2017;Picot et al., 2017;Zengin et al., 2017).Amylase and α-glucosidase are two enzymes which are responsible for postprandial glucose levels. In this context, plant extracts displaying inhibitory ac-tivity against these enzymes are being investigated as novel therapeutic target for the treatment of diabetes mellitus (Agarwal and Gupta, 2016; Van De Laar et al., 2005). In the leaf extracts, the ethyl acetate extract showed the highestα-amylase inhibition (0.84 mmol ACAE/g) while among the stem bark extracts, the methanol extract was the most ef-fective inhibitor (0.79 mmol ACAE/g). Inα-glucosidase inhibition, the best inhibition was provided by the methanol extract of the leaf (6.13 mmol ACAE/g) when compared with other solvents. In the stem bark extracts, the ethyl acetate one was the most effective α-glucosidase inhibitor (5.25 mmol ACAE/g). The stem bark methanolic extract was not active onα-glucosidase.
Another key enzyme is tyrosinase, which is involved in the catalysis of the first and only rate-limiting step in melanogenesis, a process which is responsible for the formation of the pigment melanin in skin.
Fig. 2. Multivariate analysis; (A: unsupervised principal component (PCA) in according to biological activities; B: hierarchical cluster analysis (heat map) based on biological activities; C: Sparse Partial Least Squares Discriminant Analysis (sPLS-DA) results obtained from biological activities of 6 extracts; D: biological activities outlining the difference between the two parts and the weight of all studied bioactivities on the first component of sPLS-DA; E: correlation between total bioactive compounds and biological activities).
Table 2
Antioxidant activities of the tested extracts.
Parts-Solvents DPPH (mg TE/g extract) ABTS (mg TE/g extract) CUPRAC (mg TE/g extract) FRAP (mg TE/g extract) Phosphomolybdenum (mmol TE/g)
Metal chelating ability (mg EDTAE/g) Leaves-EA 38.08 ± 1.09d 73.15 ± 1.06e 172.86 ± 2.19e 60.09 ± 3.28e 2.53 ± 0.19c 45.49 ± 1.42a Leaves-MeOH 72.98 ± 0.09a 440.32 ± 9.85b 511.48 ± 6.90b 352.96 ± 8.08b 3.35 ± 0.14b 33.01 ± 0.52b Leaves-Water 67.24 ± 2.08b 233.61 ± 3.76c 350.18 ± 5.00d 214.11 ± 2.38d 1.98 ± 0.07d 19.93 ± .44c Stem Bark-EA 51.12 ± 0.78c 84.95 ± 2.00d 137.09 ± 2.81f 66.19 ± 1.91e 1.73 ± 0.03e na Stem Bark-MeOH 73.23 ± 0.01a 490.71 ± 3.26a 717.16 ± 18.96a 440.30 ± 2.76a 4.00 ± 0.16a 5.62 ± 0.73e Stem Bark -Water 68.74 ± 0.29b 231.05 ± 1.51c 418.40 ± 9,88c 231.71 ± 5.03c 2.73 ± 0.13c 10.94 ± 0.66d
*Values expressed are means ± S.D. of three parallel measurements. TE: Trolox equivalent; EDTAE: EDTA equivalent; na: not active; EA: Ethyl acetate; MeOH:
From this point, to manage hyperpigmentation, there is an increasing demand for the downregulation of melanogenesis through the inhibi-tion of tyrosinase (Chang, 2012). In the solvent extracts of the leaf of C. perpulchrum, the best inhibitory effect for tyrosinase was provided by methanol one (159.10 mg KAE/g) followed by the ethyl acetate and aqueous. Similar order was also reported for the stem bark extracts.
Additionally, the results showed that the extracts possessed the highest level of TPC and TFC (methanol extract of stem bark and leaf, respectively) showed high enzyme inhibition against AChE, BChE, tyrosinase,α-amylase, and α-glucosidase (only the leaf methanol ex-tract for the latter), which indicate that phenolic compounds act as enzyme inhibitors. Furthermore, HPLC-ESI-MSn analysis revealed a number of bioactive compounds which could contribute to the enzyme inhibitory properties of the extracts. For instance, (Gocer et al., 2016) found that taxifolin displayed a non-competitive inhibition on AChE with IC50value of 30.1 nM. Taxifolin also non-competitively inhibited α-glucosidase (Ki= 347μM) compared to the competitive inhibition exerted by quercetin (Ki= 6.8 μM) (Proença et al., 2017). The in-hibitory effect of quercetin against AChE, BChE, α-amylase, and tyr-osinase, was also observed by (Fan et al., 2017; Jhong et al., 2015; Proença et al., 2017;Szwajgier, 2014), who found that quercetin in-hibited tyrosinase, and the inhibition occurred in a reversible and competitive manner. Also, quercetin could be chelated the copper in the active site of tyrosinase and thus it could be inhibited.
3.6. Antimicrobial evaluation of methanolic extracts
Antimicrobial activity analysis was performed using methanolic extracts of leaf and stem bark of C. perpulchrum on twelve test bacteria by MIC method (Table 4). According to the results, all tested bacteria were inhibited by methanolic extracts of C. perpulchrum. MIC values were ranged between 62.5-125μg/ml. Stem bark extracts of C. perpul-chrum was very effective on Salmonella kentucky, Proteus vulgaris and Staphylococcus aureus ATCC 25923 at low concentrations (62.5μg/ml). Besides, Enterobacter aerogenes ATCC 13048 and Serratia marrescens were inhibited by leaf extracts of C. perpulchrum with the concentration of 62.5μg/ml. MBC values were between 125-500 μg/ml and 250-500μg/ml for stem bark and leaf samples, respectively. In addition, the lowest MBC value belonged to stem bark extract which affected with P. vulgaris with the MBC value of 125μg/ml. It can be concluded that leaf and stem bark extracts were very effective on tested Gram negative and Gram positive bacteria and mainly on Gram negative bacteria at low concentrations. P. vulgaris is a facultative anaerobe which causes sev-eral infections including wound, respiratory tract and bloodstream (Kim et al., 2003;Stock, 2003). The range of antibiotics which are ef-fective on P. vulgaris has been limited and drug resistance may be de-veloped in inappropriate usage of antibiotics (Luzzaro et al., 2001;Sen et al., 2019). Methanolic stem bark extract of C. perpulchrum can be an alternative for the treatment of P. vulgaris with the lower MBC value.
Especially, Chrysophyllum albidium extracts including leaf, fruit, seed, stem and root barks and essential oil were tested for antimicrobial activity in previous studies. Variable extracts of all these parts of C.
albidium was effective in different levels on tested bacteria develop-ment. Among them, inhibition of root and seed crude extracts of C. albidium on Bacillus subtilis (Okoli and Okere, 2010), essential oil of C. albidum seed on Salmonella typhi, (Osuntokun et al., 2017), leaf, seeds and fruits extracts of C. albidum on methicillin resistant Staphylococcus aureus (George et al., 2018) stand out prominently. These antimicrobial effects were attributed to the phenolic ingredients in C. albidium (Okoli and Okere, 2010). In the current study, antimicrobial activity and lower MBC values could be explained by the rich phenolic content of stem bark of C. perpulchrum. Besides, this study is one of the limited studies which is improving our knowledge about antimicrobial activities of C. perpulchrum in the Chrysophyllum genus.
3.7. Cytotoxic evaluation of methanol extracts
Recently, plant extracts and bioactive compounds from plants pro-mise hopeful development in the treatment of cancer; our research group has also determined cytotoxic effects of many plant extracts be-fore (Mahomoodally et al., 2019; Okoli and Okere, 2010; Locatelli et al., 2018;Uysal et al., 2018;Yerlikaya et al., 2017). The methanol extracts of leaf and stem bark of C. perpulchrum were evaluated for cytotoxic activity in the current study. HeLa cells (cervical cancer cell line) were treated with variable doses (62.5, 125, 250, 500, 1000μg/ ml) of C. perpulchrum methanol extracts from leaf and stem bark sam-ples for 48 h (Figs. 3 and 4). In addition, control cells were treated with 0.1% DMSO. MTT test was performed to determine the viability of the cancer cells against extracts. Based on the MTT results, C. perpulchrum stem bark extracts displayed the highest prevention against to HeLa cell
Table 3
Enzyme inhibitory properties of the tested extracts.
Parts-Solvents AChE (mg GALAE/g extract)
BChE (mg GALAE/g extract)
Tyrosinase (mg KAE/g extract)
α-Amylase (mmol ACAE/g extract)
α-Glucosidase (mmol ACAE/g extract) Leaves-EA 4.59 ± 0.10c 0.52 ± 0.04d 114.97 ± 1.68d 0.84 ± 0.03a 5.67 ± 0.35b Leaves-MeOH 4.90 ± 0.05b 0.69 ± 0.02c 159.10 ± 0.16a 0.82 ± 0.04a 6.13 ± 0.01a Leaves-Water 1.09 ± 0.12d 0.17 ± 0.06f 65.05 ± 1.34e 0.14 ± 0.01c 5.64 ± 0.05c Stem Bark-EA 4.99 ± 0.03b 0.77 ± 0.03b 120.90 ± 0.92c 0.69 ± 0.03b 5.25 ± 0.05b Stem Bark-MeOH 5.21 ± 0.01a 0.94 ± 0.01a 152.58 ± 0.58b 0.79 ± 0.04a na Stem Bark-Water 1.10 ± 0.16d 0.24 ± 0.03e 36.88 ± 0.97f 0.16 ± 0.01c 4.92 ± 0.05d
*Values expressed are means ± S.D. of three parallel measurements. GALAE: Galatamine equivalent; KAE: Kojic acid equivalent; ACAE: Acarbose equivalent; na: not
active; EA: Ethyl acetate; MeOH: Methanol. Different letters indicate differences in the extracts (p < 0.05). Table 4
Antimicrobial evaluation of C. perpulchrum stem bark and leaves extracts and determination of their MBC and MIC values against various Gram positive and Gram negative bacteria strains.
Microorganisms Stem bark
-Leaves Extracts, MBC Stem bark -Leaves Extracts, MIC Ciproflaxin – Standard drug Enterobacter aerogenes ATCC 13048 250 - 250 62.5-125 50 Salmonella kentucky 250 - 500 62.5– 125 50 Enterococcus durans+ 250 - 500 125– 125 50 Listeria monocytogenes ATCC 7644 250 - 500 125– 125 50 Pseudomonas aeroginosa 250 - 500 125– 125 50 Enterococcus faecium+ 250 - 250 125– 125 50 Alfa streptococcus haemolyticus+ 250 - 250 125– 125 50 Staphylococcus epidermis+ 500 - 250 125– 125 50 Serratia marrescens 250 - 250 62.5-125 50 E. coli 250 - 250 125– 125 50 Proteus vulgaris 125 - 500 62.5– 125 50 Staphylococcus aureus ATCC 25923+ 250 - 500 62.5– 125 50
MBC: minimal bactericidal concentration (μg/ml); MIC: minimal inhibition concentration (μg/ml);+Gram positive.
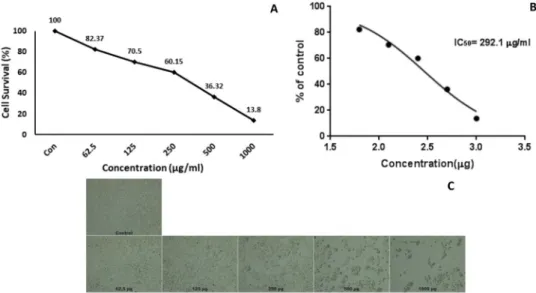
line with the IC50value of 264μg/mL after 48 h. In addition, after 48 h treatment, leaf extract of C. perpulchrum showed inhibition against the HeLa cell line with the IC50value of 292.1μg/mL. Cytotoxicity of ex-tracts on HeLa cells increased when higher concentrations applied and it can be said that the effect of C. perpulchrum extracts might be in a dose dependent manner. In addition, cytotoxic activity of stem bark samples in lower concentration than those of leaf extracts may be at-tributed to the rich phenolic content of methanol extracts of bark samples.
When considering previous studies regarding cytotoxic effects of Chrysophyllum genius on cancer cells, there have been very few studies, which included Chrysophyllum oliviforme and Chrysophyllum cainito species. High concentrations of ethyl acetate fraction of ethanolic ex-tract of C. cainito fruits [25–200 GAE (Gallic acid equivalents) μg/mL] displayed significant relation with cell death with the EC50value of 133 GAEμg/mL on osteosarcoma cells (Li et al., 2015). In another study, HeLa cells were treated with different concentrations of synthesized silver nanoparticles of satin leaf (C. oliviforme) extract. According to the results, silver nanoparticles significantly inhibited Hela cells
proliferation with an IC50value of 40.365μg/ml, in a dose dependent manner (Varghese et al., 2015). These studies are consistent with the current study for the positive correlation between dose and inhibition on cancer cells. Moreover, other studies revealed the preventive role of members of Chrysophyllum genus. For example, the total methanol ex-tract of C. perpulchrum decreased the blood sugar and normalized blood pressure and other plasma constituents which previously increased in diabetic rats (Philippe et al., 2016). In addition, methanol extract of C. cainito leaves decreased the liver and kidney toxicity induced by gamma radiation in rats (Sayed et al., 2019). It has been revealed that there are also anticancer activities by the current study in addition to the pro-tective role of C. perpulchrum extracts in previous studies.
4. Conclusion
Collectively, our data represent, for thefirst time, an in-depth study of in vitro biological and polyphenolic profile of three solvent extracts of C. perpulchrum stem bark and leaf. In general, the stem bark extract showed considerable antioxidant efficiency than the leaf extracts (with
Fig. 3. MTT analysis results (A), IC50calculation (B) and Cells morphology (C) of HeLa cells which were treated with leaf extracts of C. perpulchrum at increasing
doses for 48 h. Control cells were treated with 0.1% DMSO. Data represents the means ± SE (n = 3).
Fig. 4. MTT analysis results (A), IC50calculation (B) and Cells morphology (C) of HeLa cells which were treated with stem bark extracts of C. perpulchrum at
the exception of the metal chelating effect). The cholinesterase in-hibitory property of the stem bark extract was also higher than the leaf while the latter was generally the most effective amylase and α-glucosidase inhibitors. In addition, methanol stem bark extract had a strong antimicrobial and anticancer activities, as well. Over viewing the biological properties mentioned above, the use of C. perpulchrum could enables a certain level of health protection against oxidative damages and associated chronic diseases. With the established antioxidant and enzyme inhibitory activity of this extract, the isolation of the specific active components in C. perpulchrum extract and characterization should be further investigated.
Acknowledgments
The authors gratefully acknowledge the generous technical and human support of Universidad de Jaén (ICT of Universidad de Jaén (UJA, MINECO, Junta de Andalucía, FEDER) as well asfinancial sup-port of Kastamonu University (Kastamonu University Scientific Projects Research Office (Project Number KÜBAP-01/2016-45).
References
Agarwal, P., Gupta, R., 2016. Alpha-amylase inhibition can treat diabetes mellitus. Res. Rev. J. Med. Health Sci. 5, 1–8.
Amelin, V., Bol’shakov, D., Andoralov, A., 2018. Screening and determination of pesti-cides from various classes in natural water without sample preparation by ultra HPLC–high-resolution quadrupole time-of-flight mass spectrometry. J. Anal. Chem. 73, 257–265.
Arumugam, R., Kirkan, B., Sarikurkcu, C., 2018. Phenolic profile, antioxidant and enzyme inhibitory potential of methanolic extracts from different parts of Astragalus ponticus Pall. S. Afr. J. Bot. 120, 268–273.
Bidié, A., N’guessan, B.B., Yapo, A.F., N’guessan, J., Djaman, A., 2011. Activités anti-oxydantes de dix plantes medicinales de la pharmacopée ivoirienne. Sci. Nat. 8, 1–12.
Bidie, A.D.P., Adeoti, F.M., Yapo, F.A., Tiekpa, J.W., N’guessan, J.D., Djaman, J.A., 2016. Effect of total aqueous extract of Chrysophyllum perpulchrum on hematological, bio-chemical parameters and weight growth in wistar healthy rats. Rev. Ivoir. Sci. Technol. 28, 333–348.
Bresciani, L., Calani, L., Cossu, M., Mena, P., Sayegh, M., Ray, S., Del Rio, D., 2015. (Poly) phenolic characterization of three food supplements containing 36 different fruits, vegetables and berries. Pharma Nutr. 3, 11–19.
Chang, T.-S., 2012. Natural melanogenesis inhibitors acting through the down-regulation of tyrosinase activity. Materials 5, 1661–1685.
Choi, C.W., Song, S.B., Oh, J.S., Kim, Y.H., 2015. Antiproliferation effects of selected Tanzania plants. Afr. J. Tradit. Complement. Altern. Med. 12, 96–102.
Deng, Y., Zhao, Y., Padilla-Zakour, O., Yang, G., 2015. Polyphenols, antioxidant and antimicrobial activities of leaf and bark extracts of Solidago canadensis L. Ind. Crops Prod. 74, 803–809.
Dewan, S., 2018. Global Markets for Enzyme Inhibitors. BCC research (Accessed 22 July 2018). https://www.bccresearch.com/market-research/biotechnology/global-markets-for-enzyme-inhibitors-report-bio057c.html.
Fan, M., Zhang, G., Hu, X., Xu, X., Gong, D., 2017. Quercetin as a tyrosinase inhibitor: inhibitory activity, conformational change and mechanism. Food Res. Int. 100, 226–233.
Fischer, U.A., Carle, R., Kammerer, D.R., 2011. Identification and quantification of phe-nolic compounds from pomegranate (Punica granatum L.) peel, mesocarp, aril and differently produced juices by HPLC-DAD–ESI/MSn. Food Chem. 127, 807–821.
Friedrich, W., Eberhardt, A., Galensa, R., 2000. Investigation of proanthocyanidins by HPLC with electrospray ionization mass spectrometry. Eur. Food Res. Technol. 211, 56–64.
Future Market Insights, 2018. Enzyme Inhibitors Market: Global Industry Analysis and Opportunity Assessment 2017–2027. (Accessed 22 July 2018). https://www. futuremarketinsights.com/reports/enzyme-inhibitors-market.
George, O.A., Adenipekun, E.O., Fasogbon, S.A., Oparanozie, J.A., 2018. Antimicrobial activities of Chrysophyllum albidum leaves, fruits and seeds. Am. J. Biomed. Sci. 10, 28–44.
Gocer, H., Topal, F., Topal, M., Küçük, M., Teke, D., Gülçin,İ., Alwasel, S.H., Supuran, C.T., 2016. Acetylcholinesterase and carbonic anhydrase isoenzymes I and II in-hibition profiles of taxifolin. J. Enzyme Inhib. Med. Chem. 31, 441–447.
Gómez‐Caravaca, A.M., López‐Cobo, A., Verardo, V., Segura‐Carretero, A.,
Fernández‐Gutiérrez, A., 2016. HPLC‐DAD‐q‐TOF‐MS as a powerful platform for the determination of phenolic and other polar compounds in the edible part of mango and its by‐products (peel, seed, and seed husk). Electrophoresis 37, 1072–1084.
Grossberg, G.T., 2003. Cholinesterase inhibitors for the treatment of Alzheimer’s disease: getting on and staying on. Curr. Ther. Res. 64, 216–235.
Guédé, N.Z., N’guessan, K., Dibié, T.E., Grellier, P., 2010. Ethnopharmacological study of plants used to treat malaria, in traditional medicine, by Bete Populations of Issia (Côte d’Ivoire). J. Pharm. Sci. Res. 2, 216–227.
Huyut, Z., Beydemir,Ş., Gülçin, İ., 2017. Antioxidant and antiradical properties of se-lectedflavonoids and phenolic compounds. Biochem. Res. Int. 2017.
Hvattum, E., 2002. Determination of phenolic compounds in rose hip (Rosa canina) using liquid chromatography coupled to electrospray ionisation tandem mass spectrometry and diode‐array detection. Rapid Commun. Mass Spectrom. 16, 655–662.
Jhong, C.H., Riyaphan, J., Lin, S.H., Chia, Y.C., Weng, C.F., 2015. Screening alpha‐glu-cosidase and alpha‐amylase inhibitors from natural compounds by molecular docking in silico. BioFactors 41, 242–251.
Kajdžanoska, M., Gjamovski, V., Stefova, M., 2010. HPLC-DAD-ESI-MSn identification of phenolic compounds in cultivated strawberries from Macedonia. Maced. J. Chem. Chem. Eng. 29, 181–194.
Kasote, D.M., Katyare, S.S., Hegde, M.V., Bae, H., 2015. Significance of antioxidant po-tential of plants and its relevance to therapeutic applications. Int. J. Biol. Sci. 11, 982.
Kim, B.-n., Kim, N.J., Kim, M.-n., Kim, Y.S., Woo, J.-h., Ryu, J., 2003. Bacteraemia due to tribe Proteeae: a review of 132 cases during a decade (1991–2000). Scand. J. Infect. Dis. 35, 98–103.
Lazarova, I., Zengin, G., Aktumsek, A., Gevrenova, R., Ceylan, R., Uysal, S., 2014. HPLC–DAD analysis of phenolic compounds and antioxidant properties of Asphodeline lutea roots from Bulgaria and Turkey. Ind. Crops Prod. 61, 438–441.
Lazarova, I., Zengin, G., Bender, O., Zheleva-Dimitrova, D., Uysal, S., Ceylan, R., Gevrenova, R., Aktumsek, A., Acar, M., Gunduz, M., 2015. A comparative study of Bulgarian and Turkish Asphodeline lutea root extracts: HPLC–UV profiles, enzyme inhibitory potentials and anti-proliferative activities against MCF-7 and MCF-10A cell lines. J. Funct. Foods 15, 254–263.
Li, L.-B., Lin, S., Yan, J., Wang, Q.-L., Fan, Z.-Y., Dong, Q.-R., Qin, J.-Z., Xie, Z.-G., 2015. Poly-phenolic fraction of Chrysophyllum cainito extract induces cell death in osteo-sarcoma cells. Bangladesh J. Pharmacol. 10, 972–979.
Llorent-Martínez, E.J., Zengin, G., Fernández-de Córdova, M.L., Bender, O., Atalay, A., Ceylan, R., Mollica, A., Mocan, A., Uysal, S., Guler, G.O., 2017. Traditionally used Lathyrus species: phytochemical composition, antioxidant activity, enzyme inhibitory properties, cytotoxic effects, and in silico studies of L. czeczottianus and L. nissolia. Front. Pharmacol. 8, 83.
Locatelli, M., Yerlikaya, S., Baloglu, M.C., Zengin, G., Altunoglu, Y.C., Cacciagrano, F., Campestre, C., Mahomoodally, M.F., Mollica, A., 2018. Investigations into the ther-apeutic potential of Asphodeline liburnica roots: in vitro and in silico biochemical and toxicological perspectives. Food Chem. Toxicol. 120, 172–182.
Luzzaro, F., Perilli, M., Amicosante, G., Lombardi, G., Belloni, R., Zollo, A., Bianchi, C., Toniolo, A., 2001. Properties of multidrug-resistant, ESBL-producing Proteus mirabilis isolates and possible role ofβ-lactam/β-lactamase inhibitor combinations. Int. J. Antimicrob. Agents 17, 131–135.
Mahomoodally, M.F., 2013. Traditional medicines in Africa: an appraisal of ten potent African medicinal plants. Evid. Based Complementary Altern. Med. 2013.
Mahomoodally, M.F., Yerlikaya, S., Llorent-Martínez, E.J., Uğurlu, A., Baloglu, M.C., Altunoglu, Y.C., Mollica, A., Dardenne, K.K., Aumeeruddy, M.Z., Puchooa, D., 2019. Pharmacological and polyphenolic profiles of Phyllanthus phillyreifolius var. commer-sonii Müll. Arg: an unexplored endemic species from Mauritius. Food Res. Int. 115, 425–438.
Mämmelä, P., 2001. Phenolics in selected European hardwood species by liquid chro-matography–electrospray ionisation mass spectrometry. Analyst 126, 1535–1538.
Mena, P., Calani, L., Dall’Asta, C., Galaverna, G., García-Viguera, C., Bruni, R., Crozier, A., Del Rio, D., 2012. Rapid and comprehensive evaluation of (poly) phenolic com-pounds in pomegranate (Punica granatum L.) juice by UHPLC-MSn. Molecules 17, 14821–14840.
Okoli, B., Okere, O., 2010. Antimicrobial activity of the phytochemical constituents of Chrysophyllum albidum G. Don_Holl.(African Star apple) plant. J. Res. Natl. Dev. 8, 1035–1037.
Osuntokun, O.T., Ayegbusi, B., Yusuf-Babatunde, A., Ige, O., 2017. Assessment of anti-microbial, phytochemical screening and gas chromatography-mass spectro-photometric profile of crude Chrysophyllum albidum essential oil. Chem. Res. J. 2, 68–85.
Ozgen, S., Kilinc, O.K., Selamoğlu, Z., 2016. Antioxidant activity of quercetin: a me-chanistic review. Turk. J. Agric. Food Sci. Technol. 4, 1134–1138.
Phatak, R.S., Hendre, A.S., 2014. Total antioxidant capacity (TAC) of fresh leaves of Kalanchoe pinnata. J. Pharmacogn. Phytochem. 2, 32–35.
Philippe, B.A., Karine, N., Barthélemy, A.K., Noél, Z.G., Joseph, D.A., Hosttetmann, K., 2010. Bio-guided isolation of antioxidant compounds from Chrysophyllum perpul-chrum, a plant used in the Ivory Coast pharmacopeia. Molecules 15, 6386–6398.
Philippe, B.A.D., Justine, T.W., Brice, B.K., 2016. Effect of total methanolic extract of Chrysophyllum perpulchrum on lipemia and blood pressure in rats made diabetic by streptozotocin. Int. J. Biochem. Res. Rev. 13, 1–9.
Picot, M.C., Zengin, G., Mollica, A., Stefanucci, A., Carradori, S., Mahomoodally, M., 2017. In vitro and in silico studies of mangiferin from Aphloia theiformis on key en-zymes linked to diabetes type 2 and associated complications. Med. Chem. 13 (7), 633–640.
Plumb, G., de Pascual-Teresa, S., Santos-Buelga, C., Rivas-Gonzalo, J.C., Williamson, G., 2002. Antioxidant properties of gallocatechin and prodelphinidins from pomegranate peel. Redox Rep. 7, 41–46.
Proença, C., Freitas, M., Ribeiro, D., Oliveira, E.F., Sousa, J.L., Tomé, S.M., Ramos, M.J., Silva, A.M., Fernandes, P.A., Fernandes, E., 2017.α-Glucosidase inhibition by fla-vonoids: an in vitro and in silico structure–activity relationship study. J. Enzyme Inhib. Med. Chem. 32, 1216–1228.
Sayed, D., Nada, A., Mohamed, M.A.E.H., Ibrahim, M., 2019. Modulatory effects of Chrysophyllum cainito L. extract on gamma radiation induced oxidative stress in rats. Biomed. Pharmacother. 111, 613–623.
Sen, F., Ganim, M.A., Baloglu, M.C., Aygun, A., Sayiner, H.S., Altunoglu, Y.C., Kandemirli, F., Demirkan, B., Kuyuldar, E., Bulut, E., 2019. Synergistic and antagonistic effects of phenylalanine and various antibiotics on the growth of pathogenic bacteria. BioNanoScience 1–7.
Shanmugam, B., Shanmugam, K.R., Ravi, S., Subbaiah, G.V., Ramakrishana, C., Mallikarjuna, K., Reddy, K.S., 2017. Exploratory studies of (-)-epicatechin, a bioactive compound of Phyllanthus niruri, on the antioxidant enzymes and oxidative stress markers in D-galactosamine-induced hepatitis in rats: a study with reference to clinical prospective. Pharmacog. Mag. 13, S56.
Stock, I., 2003. Natural antibiotic susceptibility of Proteus spp., with special reference to P. mirabilis and P. penneri strains. J. Chemother. 15, 12–26.
Szwajgier, D., 2014. Anticholinesterase activities of selected polyphenols–a short report. Pol. J. Food Nutr. Sci. 64, 59–64.
Topal, F., Nar, M., Gocer, H., Kalin, P., Kocyigit, U.M., Gülçin,İ., Alwasel, S.H., 2016. Antioxidant activity of taxifolin: an activity–structure relationship. J. Enzyme Inhib. Med. Chem. 31, 674–683.
Tropical Plants Database, 2018. tropical.theferns.info/ viewtropical.php?id=Chrysophyllum+perpulchrum.
Turan, N., Adıguzel, R., Buldurun, K., Bursal, E., 2016. Spectroscopic, thermal and anti-oxidant properties of novel mixed ligand-metal complexes obtained from sacchar-inate complexes and azo dye ligand (mnppa). Int. J. Pharmacol. 12, 92–100.
Uysal, S., Ugurlu, A., Zengin, G., Baloglu, M.C., Altunoglu, Y.C., Mollica, A., Custodio, L., Neng, N.R., Nogueira, J.M., Mahomoodally, M.F., 2018. Novel in vitro and in silico insights of the multi-biological activities and chemical composition of Bidens tripartita L. Food Chem. Toxicol. 111, 525–536.
Van De Laar, F.A., Lucassen, P.L., Akkermans, R.P., Van De Lisdonk, E.H., Rutten, G.E., Van Weel, C., 2005.α-Glucosidase inhibitors for patients with type 2 diabetes: results
from a Cochrane systematic review and meta-analysis. Diabetes Care 28, 154–163.
Varghese, R.A., Anandhi, P., Arunadevi, R., Boovisha, A., Sounthari, P., Saranya, J., Parameswari, K., Chitra, S., 2015. Satin leaf (Chrysophyllum oliviforme) extract mediated green synthesis of silver nanoparticles: antioxidant and anticancer activ-ities. J. Pharm. Sci. Res. 7, 266.
Wan, C., Yu, Y., Zhou, S., Liu, W., Tian, S., Cao, S., 2011. Antioxidant activity and free radical-scavenging capacity of Gynura divaricata leaf extracts at different tempera-tures. Pharmacog. Mag. 7, 40.
Yang, M., Shen, Q., Li, L.-Q., Huang, Y.-Q., Cheung, H.-Y., 2015. Phytochemical profiles, antioxidant activities of functional herb Abrus cantoniensis and Abrus mollis. Food Chem. 177, 304–312.
Yerlikaya, S., Zengin, G., Mollica, A., Baloglu, M.C., Celik Altunoglu, Y., Aktumsek, A., 2017. A multidirectional perspective for novel functional products: in vitro phar-macological activities and in silico studies on Ononis natrix subsp. hispanica. Front. Pharmacol. 8, 600.
Zengin, G., Ceylan, R., Katanić, J., Mollica, A., Aktumsek, A., Boroja, T., Matić, S., Mihailović, V., Stanić, S., Aumeeruddy-Elalfi, Z., 2017. Combining in vitro, in vivo and in silico approaches to evaluate nutraceutical potentials and chemical finger-prints of Moltkia aurea and Moltkia coerulea. Food Chem. Toxicol. 107, 540–553.
Zhao, P., Duan, L., Guo, L., Dou, L.-L., Dong, X., Zhou, P., Li, P., Liu, E.-H., 2015. Chemical and biological comparison of the fruit extracts of Citrus wilsonii Tanaka and Citrus medica L. Food Chem. 173, 54–60.