Yoğun Bakım Hastalarından İzole Edilen Enterokok İzolatlarında Çeşitli Virulans Ve Direnç Genlerinin Varlığının Araştırılması
Tam metin
Şekil
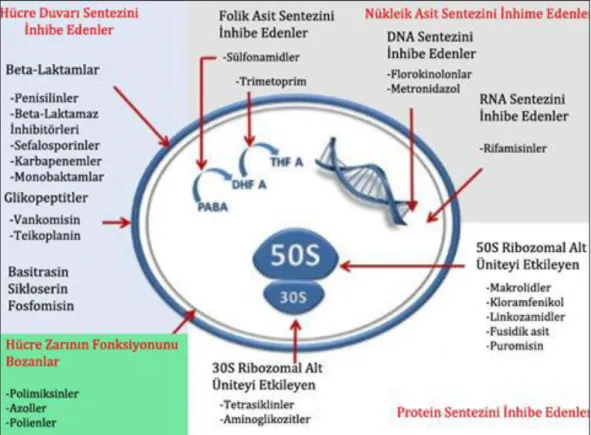
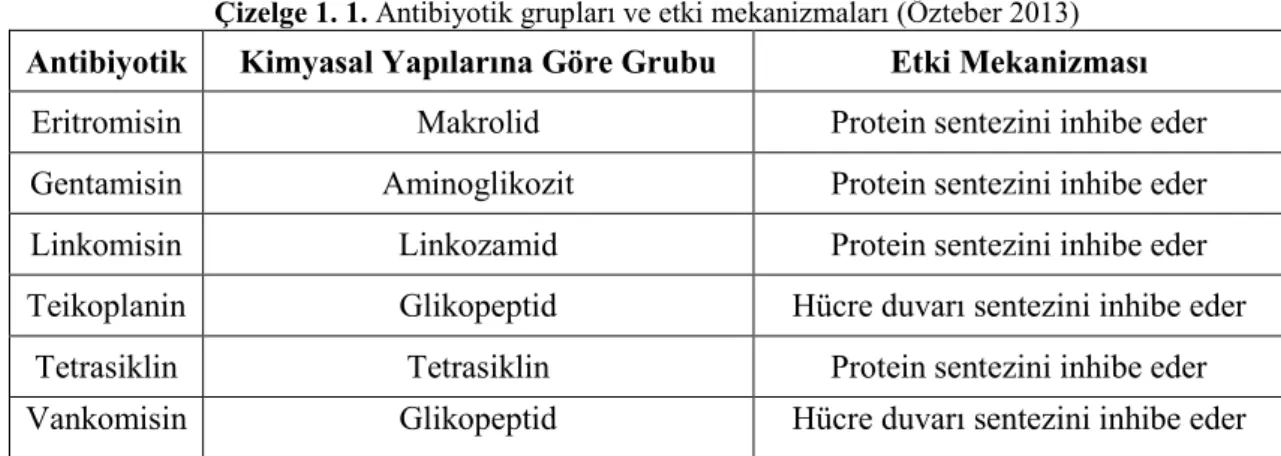

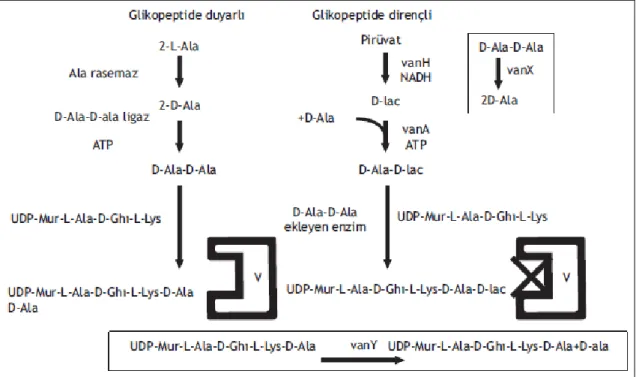
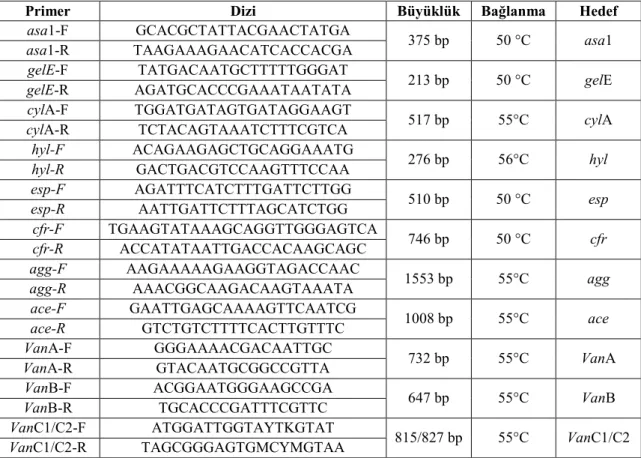

Outline
Benzer Belgeler
Giriş: Bu çalışmada, Diyarbakır Eğitim ve Araştırma Hastanesi, Anestezi Yoğun Bakım ve Reanimasyon Klini- ği’nden gelen hasta örneklerinden soyutlanan maya man-
ABSTRACT Objective: In this study, it was aimed to determine vancomycin resistance mechanisms, virulence genes and clonal relationships of 23 vancomycin-resistant Enterococcus
Çalışmamızda Karadeniz Teknik Üniversitesi Tıp Fakültesi Farabi Hastanesi Tıbbi Mikro- biyoloji Laboratuvarında, Ekim 2016-Eylül 2018 tarihleri arasında çeşitli
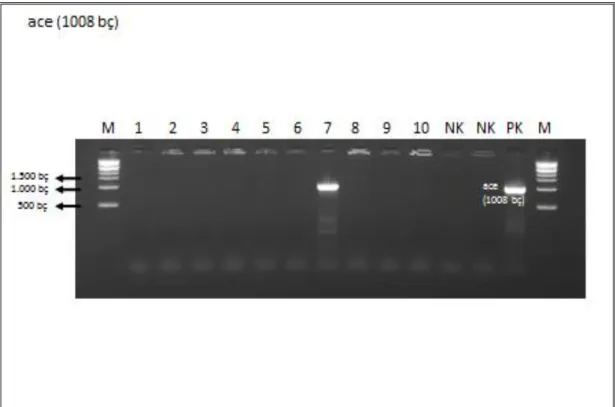
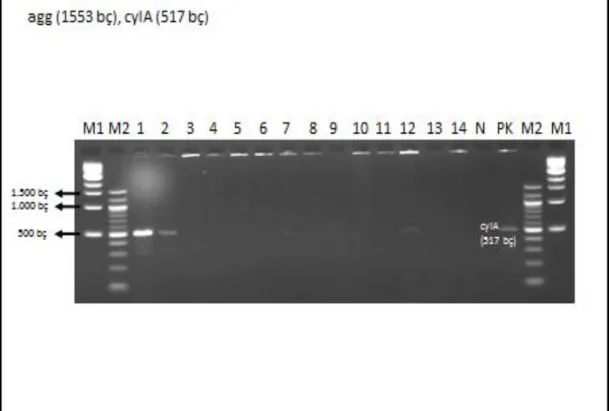
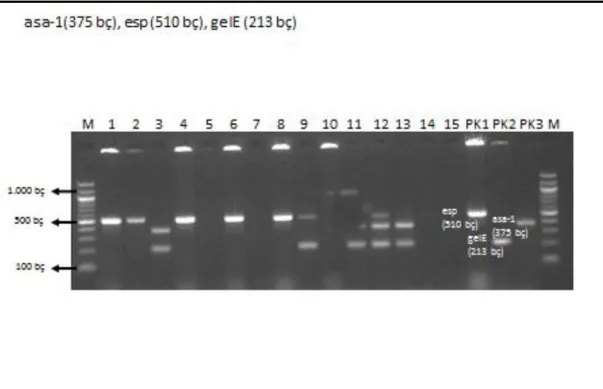
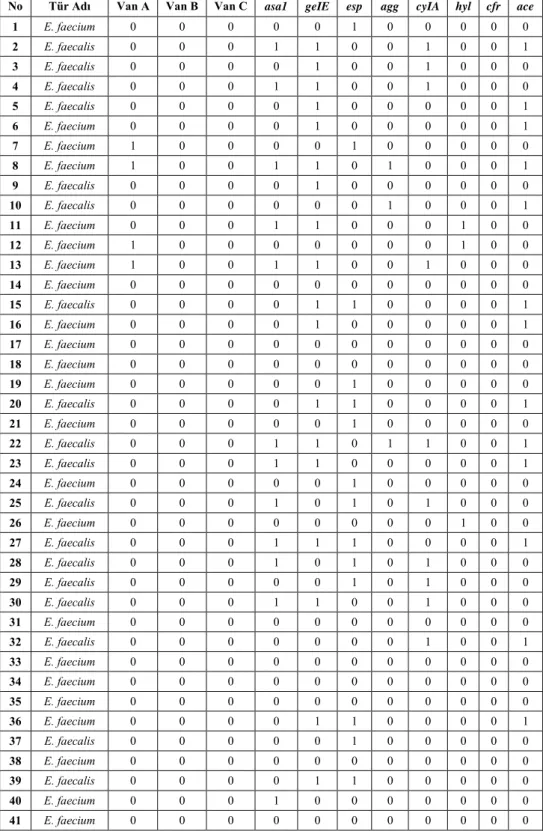
Vankomisin dirençli ve duyarlı izolatların virulans faktörleri karşılaştırıldığında; vankomisin dirençli enterokok ( VRE) E.faecium izolatlarında esp gen düzeyi
Geriye kalan beş olguda doğrudan ve daha önce rektal kolonizasyonu olan dört olguda da sonradan olmak üzere toplam dokuz olguda enfeksiyon (beş bakteriyemi, üç kateter
Zeynep Kamil Kadın ve Çocuk Hastalıkları Eğitim ve Araştırma Hastanesi Yenidoğan YBÜ’de 11 Ağustos 2009 tarihinde (birinci gün) Bartter sendromu tanısıyla takip
Sonuç olarak, nadiren de olsa Leuconostoc türlerinin insanlarda menenjit etkeni olabi- leceği ve gram-pozitif koklarda vankomisin direncinin saptanması halinde Leuconostoc
Aminoglikozidler ve beta-laktamlar arasında in vitro sinerji, vahşi tip ve çoklu ilaç direncine sahip izolatlar da dahil olmak üzere hem Gram negatif hem de
