Improving Nutritional Qualities of Tomato Pomace by Pleurotus ostreatus and Phanerochaete
chrysosporium Fermentation
Sülhattin YAŞAR1, Ramazan TOSUN2
Igdır University, Faculty of Agriculture Department of Animal Science, 76000,
1https://orcid.org/0000-0001-9334-1303, 2https://orcid.org/0000-0002-8209-6362
ABSTRACT
In this study, it was aimed to improve nutrient quality by fermenting tomato pomace with Pleurotus ostreatus (P. ostreatus) and Phanerochaete chrysosporium (P. chrysosporium). Tomato pomace was incubated for 21 days at optimized conditions of pH (3.50-5.50), temperature (24-28 oC), moisture content (68% w w-1), aeration (0,25 L min-1) and stirring rates (10 rpm). Three samples taken at each incubation time were chemically analyzed. The results indicated that fermentation with P. ostreatus and P. chrysosporium significanly increased ash content by 25 and 21%, crude protein content by 16 and 30%, respectively (P<0.05). Fermentation with P. ostreatus decreased ether extract content from 7.22% to 0.29% at 21th day (P<0.05). However, there was an increase of ether extract content with P. chrysosporium fermentation (from 7.22 to 11.62% at 21 day) (P<0.05). Crude fiber of tomato pomace with P. chrysosporium were reduced by 64% (P<0.05). Both fungal fermentations reduced total reducing sugar content by about 30% (P<0.05). Fermentation with P. ostreatus and P. chrysosporium significantly changed tannin and pectin levels (P<0.05). As a result, fungal fermentation caused to nutritionally enriched tomato pomace with added active compounds, and could be used as functional feed in animal nutrition.
Research Article Article History Received : 25.05.2019 Accepted : 31.10.2019 Keywords Functional feed Phanerochaete chrysosporium Pleurotus ostreatus
Solid state fermentation Tomato pomace
Domates Posasının Pleurotus ostreatus ve Phanerochaete chrysosporium Fermentasyonu ile Besleyici
Değerinin Artırılması
ÖZET
Bu çalışmada, domates posasını Pleurotus ostreatus (P. ostreatus) ve
Phanerochaete chrysosporium (P. chrysosporium) ile fermente ederek
besin madde kalitesini artırmak amaçlanmıştır. Domates posası, optimize edilmiş pH (3.50-5.50), sıcaklık (24-28 oC), nem içeriği (%68), havalandırma (0.25 L dk-1) ve karıştırma hızı (10 rpm) koşullarında 21 gün inkubasyona bırakılmıştır. Fungal fermentasyon sonuçlarına göre ham kül içeriği %21-25 oranında ve ham protein içeriği ise %16-30 oranında artmıştır (P<0.05). P. ostreatus ile fermentasyonda, 21. günde ham yağ içeriği %7.22’den %0.29’a önemli derecede azalmıştır (P<0.05). Ancak ham yağ içeriği P. chrysosporium ile yürütülen fermentasyonda artmıştır (%7.22’den %11.62’e kadar artış) (P<0.05).
P. chrysosporium ile yürütülen fermentasyonda ham selüloz içeriği
yaklaşık %64 oranında azalmıştır (P<0.05). Domates posasının her iki fungal fermentasyonunda da toplam redükte şeker içeriği yaklaşık %30’a kadar düşmüştür (P<0.05). Tanin ve pektin içeriği her iki fungal fermentasyonunda da önemli derecede değişmiştir (P<0.05). Sonuç olarak, fungal fermentasyonu domates posasının besleyici değerinin artmasına, biyolojik olarak fonksiyonel bileşiklerin oluşmasına sebep olmuştur.
Araştırma Makalesi Makale Tarihçesi Geliş Tarihi : 25.05.2019 Kabul Tarihi : 31.10.2019 Anahtar Kelimeler İşlevsel yem Phanerochaete chrysosporium Pleurotus ostreatus
Katı faz fermentasyon Domates posası
To Cite : Yaşar S, Tosun S 2020. Improving Nutritional Qualities of Tomato Pomace by Pleurotus ostreatus and
Phanerochaete chrysosporium Fermentation. KSU J. Agric Nat 23 (2): 527-534. DOI: 10.18016/
ksutarimdoga.vi.629347
INTRODUCTION utilisation of agro-industrial waste products causing enviromental pollution to a greater extend. Tomato
pomace is a by-product of tomato processing industry, and composed of tomato skin, seed and pulp. The world’s annual tomato waste production reaches up to 11 million ton per year including 4 million tons of tomato pomace (FAO, 2016). Tomato pomace contains appreciable amount of proteins, lipids, carbohydrates, amino acids, carotenoids and minerals (Frexio et al., 2012; Liu et al., 2013; Ergun and Urek, 2017; Waldbauer et al., 2017; Ulker et al., 2018). It has mainly been used as feed material or soil fertilizer (Knoblich et al., 2015; Bennamoun et al., 2016). Tomato pomace can directly be fed to ruminant animals as fresh or dried forms and as a part of silage at appreciable amounts. The ruminant animals can easily utilize from the nutrients of tomato pomace as a result of microbial digestion of the rumen (Weiss et al., 1997; Mirzaei-Aghsaghali and Maheri-Sis, 2008; Ziaei and Molaei, 2010; Abdollahzadeh et al., 2010). In contrary, poultry species can not completely utilize from tomato pomace so as to ruminant animals since the digestive tract of most of the poultry species, especially young growing birds, do not sufficiently secret specific enzymes degrading the nutrients such as crude fibre (CF), tannin and pectin which are mostly considered as antinutritional factors (ANFs) (King and Zeidler, 2004; Al-Betawi, 2005; Wadhwa and Bakshi 2016; Yasar and Tosun, 2019). In addition, tomato pomace is a seasonal product and not available throughout the entire year and difficult for conversation due to its high moisture content of 75%. Moreover, drying tomato pomace at commercial scale to produce animal feed has been found not economically feasible (Weiss et al., 1997), but there are novel processing treatments including drying to produce bioactive compounds such as antioxidants, lycopene, oils and protein as food and feed ingredients (Lu et al., 2019).
Tomato pomace has been successfully used as fermentation substrate by the industry for the production of functional bioactive molecules (enzymes, organic acids, aromatic compounds and antimicrobial agents), for improvement of its nutritional qualities and for reduction of its contents of ANFs (Raimbault, 1998; Singhaniaa et al., 2009; Özşölen, 2010; Afşin, 2010; Kurt and Buyukalaca, 2010; Ravichandran and Vimala, 2012; Mukherjee et al., 2016). Solid state fermentation (SSF) is defined as fermentation of insoluble solid substrates immersed in free water by microorganism cultures similar to their natural environment (Afşin, 2010). For instance, several biologically active enzymes inlcuding cellulases and oxidases (Verma and Madamwar, 2002; Rashad et al., 2009; Iandolo et al., 2011; Yoon et al., 2014; Bennamoun et al., 2016; Ergun and Urek, 2017) are produced by the P. ostreatus and Phanerochaete
chrysosporium (P. chrysosporium) fermentations. P.
chrysosporium was considered an ideal fungal
microorganism in order to increase the levels of phenolic compounds of apple fruit pomace (Ajila et al., 2011; Yasar and Tosun, 2018).
The fungals of Rhizopus stolonifer LAU 07, Candida
utilis, Trichoderma viride, Aspergillus niger,
Fusarium, P. ostreatus and P. chrysosporium
(Villas-Boas et al., 2003; Lateef et al., 2008; Yasar and Tosun, 2018), the yeast of Saccharomyces cervisiae and bacteria of Bacillus subtilis (Azza et al., 2013) have been used in SSF processes of agricultural by-products for crude protein enrichment. In addition, fungal microorganisms was reported to break down cellulose, hemicellulose and other complex polysaccharides in industrial by-products (Rashad et al., 2009; Díaz-Godínez et al., 2012). Previously studies reported that fermenting tomato pomace with several microorganisms increased the amount of ash and crude protein (CP) and decreased the levels of CF and hemicellulose (Assi and King, 2008; Azza et al., 2013; Roja et al., 2017; Yasar and Tosun, 2019).
Having evaluated all the above stated results, P.
ostreatus and P. chrysosporium could ideally act as
best fungal microorganisms to nutritional enrichment of tomato pomace at optimum conditions of 4.0–5.0 of pH, 25-35 oC of temperature and low stirring rates at occational intervals, which were selected from the literature and optimized and controlled throughout the study using a modern bioreactor. Therefore, the objective of this study was to test the effect of two fungal microorganisms used in SSF on the changes in nutritional composition of pomace.
MATERIALS and METHODS
Tomato pomace optained from a local provider dried and ground to pass a sieve with 3 mm were supplemented with nutrients as shown in Table 1 and was further autoclaved at 120 ℃ for 15 min. Two fungal microorganisms, Pleurotus ostreatus (P. ostreatus)
and Phanerochaete chrysosporium (P. chrysosporium)
obtained from DSMZ (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Germany) were cultivated according to the supplier instruction to collect inoculating spores. Optimum fermentation conditions fixed in the study (Table 1) were selected from the literature and optimized by using a laboratory bioreactor of 2.5-3 L working capacity, LabforEtOlH 5 (Infors Ltd., Switzerland). A blank fermentation experiment was conducted with no fungal inoculation. The fixed pH values were well optimized by peristaltic pumps using buffer solutions of 0.1 M sodium acetate (pH=1.5) and 0.1 M sodium bicarbonate (pH=9.75). At each sampling period (days), three independent samples were taken from each of fermentation experiments (Table 1) were consequently analysed three times as replicates for the determinations of nutritional and antinutritional factors parameters.
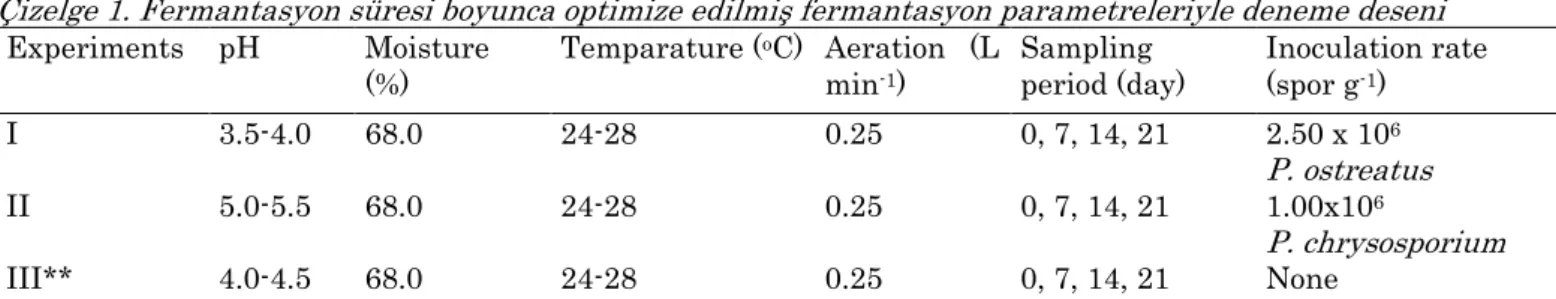
Table 1. Experimental design with optimised fermentation parameters fixed throughout the fermentation period
Çizelge 1. Fermantasyon süresi boyunca optimize edilmiş fermantasyon parametreleriyle deneme deseni
Experiments pH Moisture
(%) Temparature (
oC) Aeration (L min-1)
Sampling
period (day) Inoculation rate (spor g-1)
I 3.5-4.0 68.0 24-28 0.25 0, 7, 14, 21 2.50 x 106
P. ostreatus
II 5.0-5.5 68.0 24-28 0.25 0, 7, 14, 21 1.00x106
P. chrysosporium
III** 4.0-4.5 68.0 24-28 0.25 0, 7, 14, 21 None
*Added nutrients in experiment I, II and III were 20 g (NH4)2SO4, 10 g NH4Cl, 10 g CH4N2O and 60 g Molasses, and there was a constant rate of stirring rate (10 rpm for 2 min at every 12 h). **Blank fermentation, a non-pH optimised experiment (initial pH of 4.00 did not change throughout the fermentation period) under sterile fermentation conditions, the same as in experiments I and II
Thus, 9 independent replicates per treatment were obtained, and data was analysed according to a general linear model (GLM) of variance analysis, where the differences between the treatments were separated at 0.05 significance level using SPSS software (IBM SPSS Statistics 22.0 for Windows). Fungal growth was determined by the method of TS ISO 21527-2: 2008 and the contents of dry matter (DM, %), ash %, crude protein (CP, %), crude fiber (CF, %), ether extract (EE, %) and reducing sugar (RS, %) by the methods specifically expressed in AOAC (2005). Pectin (Wang and Zhang, 1999) and tannin (Chemesova and
Chizhikov, 2004) were spectrophotometrically analysed.
RESULT and DISCUSSION
The results of Table 2 that the growth rate of P.
ostreatus in experiment I and P. chrysosporium in
experiment II significantly increased by 3 log at the end of 21 days of fermentation (P<0.05). And the pH was well controlled in the pre-fixed ranges in both experiments (Table 1), indicating a successful fermentation of fungal microorganism on tomato pomace.
Table 2. Fungal growth rate and pH of fermenting substrate at 0, 7, 14 and 21 days of fermentation
Çizelge 2. 0, 7, 14 ve 21 günlük fermantasyonda fungal gelişim oranı ve fermente substratın pH değeri
Days cfu g-1 pH
Experiments-I
(P. ostreatus) Experiments-II (P.chrysosporium) Experiments III (Control) Experiments I(P.ostreatus) Experiments II (P.chrysosporium) Experiments III (Control) 0 7.9x105±0.05d 3.07x105±0.05c 0.0±0.05 4.05±0.20 5.50±0.10 4.00±0.10 7 8.0x107±0.10b 1.7x108±0.20a 0.0±0.10 3.70±0.20 5.30±0.15 4.00±0.10 14 1.3x107±0.13c 5.0x106±0.05b 0.0±0. 10 3.70±0.25 5.30±0.20 4.00±0.15 21 3.9x108±0.15a 1.3x108±0.12a 0.0±0.10 3.60±0.25 5.40±0.21 4.00±0.15 a,b,c,d Different supercripts showed significant differences between the fermentation periods at each of the coloumn parameters .
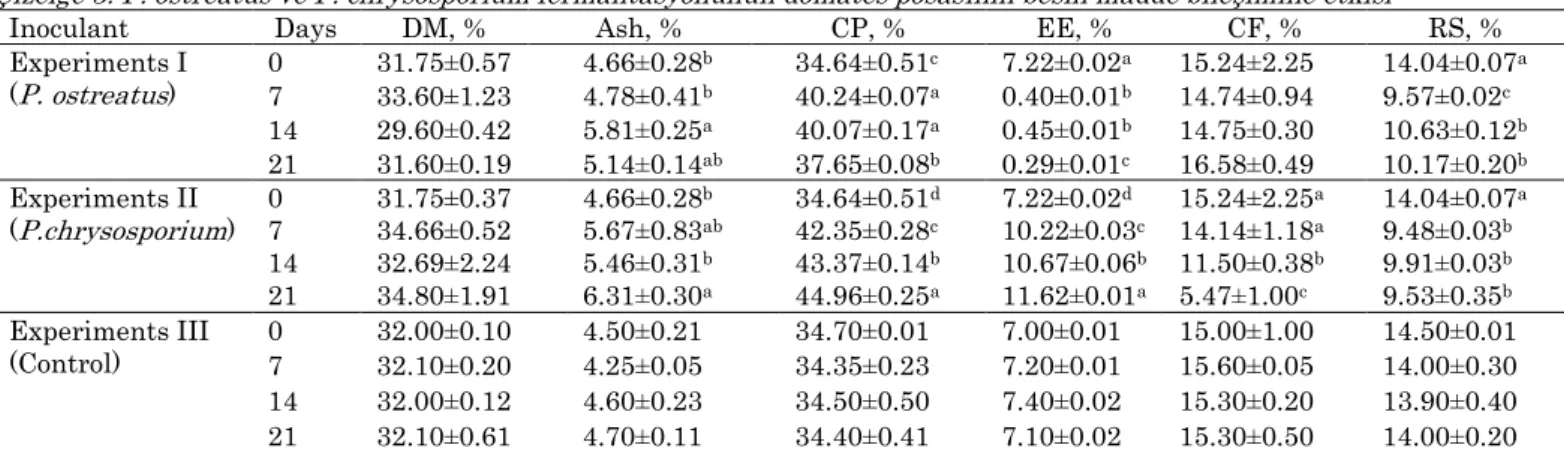
No significant changes were seen in nutritional composition of tomato pomace fermented with no fungal inoculations (P>0.05) (Table 3). Fermenting tomato pomace with two different fungal inoculants significantly influenced its nutritional composition (P<0.05). The contents of ash significantly increased by the fermentations of both fungal species (P<0.05). The highest increases in ash content were 25% at 14 days
of P. ostreatus and 35% at 21 days of P. chrysosporium
fermentations. There were 16 and 30% increases in the CP content at the 14 days of P. ostreatus and at 21 days
of P. chrysosporium fermentations, respectively were
significant (P<0.05). The fermentation of tomato pomace with P. ostreatus significanly reduced its EE content from 7.22% at 0 day to 0.29% at 21 days. In contrary, there was a gradual increase in the EE content of tomato pomace with the P. chrysosporium
fermentation (from 7.22% at 0 day to 11.62% at 21 day). As overall there was no significant effect of P.
ostreatus fermentation on the CF content, whereas an
approximate 3-fold decrease in the CF content of tomato pomace was obtained from the P.
chrysosporium fermentation (15.24% at 0 day to 5.47%
at 21 day). The RS contents of tomato pomace were significantly reduced by about 30% in both fermentation cases (P<0.05).
Ash content of tomato pomace increased in both fungal fermentations. Dei et al. (2008) reported increased ash content of industrial by-products as a result of fungal fermentation. The reason for the inc rease in ash content was elaborated that during fermentation, microorganisms secrete enzymes which degrade complex minerals such as phosphorus in phytic acid form, and as a result of fermentation, the liberated minerals are released and thus the ash content increases. Many other studies found increased ash content of the fermenting substrates as a result of fermentation (Dei et al., 2008; Okpako et al., 2008; Aguilar et al., 2008; Altop et al., 2018).
Table 3. Nutritional composition of tomato pomace affected by P. ostreatus and P. chrysosporium fermentation Çizelge 3. P. ostreatus ve P. chrysosporium fermantasyonunun domates posasının besin madde bileşimine etkisi
Inoculant Days DM, % Ash, % CP, % EE, % CF, % RS, %
Experiments I (P. ostreatus) 0 31.75±0.57 4.66±0.28 b 34.64±0.51c 7.22±0.02a 15.24±2.25 14.04±0.07a 7 33.60±1.23 4.78±0.41b 40.24±0.07a 0.40±0.01b 14.74±0.94 9.57±0.02c 14 29.60±0.42 5.81±0.25a 40.07±0.17a 0.45±0.01b 14.75±0.30 10.63±0.12b 21 31.60±0.19 5.14±0.14ab 37.65±0.08b 0.29±0.01c 16.58±0.49 10.17±0.20b Experiments II (P.chrysosporium) 0 31.75±0.37 4.66±0.28b 34.64±0.51d 7.22±0.02d 15.24±2.25a 14.04±0.07a 7 34.66±0.52 5.67±0.83ab 42.35±0.28c 10.22±0.03c 14.14±1.18a 9.48±0.03b 14 32.69±2.24 5.46±0.31b 43.37±0.14b 10.67±0.06b 11.50±0.38b 9.91±0.03b 21 34.80±1.91 6.31±0.30a 44.96±0.25a 11.62±0.01a 5.47±1.00c 9.53±0.35b Experiments III (Control) 0 7 32.00±0.10 32.10±0.20 4.50±0.21 4.25±0.05 34.70±0.01 34.35±0.23 7.00±0.01 7.20±0.01 15.00±1.00 15.60±0.05 14.50±0.01 14.00±0.30 14 32.00±0.12 4.60±0.23 34.50±0.50 7.40±0.02 15.30±0.20 13.90±0.40 21 32.10±0.61 4.70±0.11 34.40±0.41 7.10±0.02 15.30±0.50 14.00±0.20 a,b,c, d Different supercripts showed significant differences between the fermentation periods at each of the coloumn parameters.
Lateef et al. (2008), clearly showed that fungal fermentation using the strain of Rhizopus stolonifer
LAU 07 significantly improved nutritional qualities of
some agro-wastes by increasing CP contents by 35-90% and reducing CF by 7.0 to 44% (P<0.05). Similar improvement rates were also reported by several other fungal species including P. ostreatus and P.
chrysosporium in the study of Assi and King, (2008);
Rashad et al. (2009 and 2010); Díaz-Godínez, (2012) and Yasar and Tosun, (2018). In fungal fermentation fungal growth and reproduction of micelles formed as a result of the substrate used in fermentation has been reported to increase the CP content (Altop et al., 2018). On the other hand, Oboh and Akindahunsi (2003) claimed that the increased CP content was due to the increased enzymes which are in nitrogenous nature. It is thought that the increase of CP content in the fermentation of tomato pomace with P. ostreatus and
P. chrysosporium is due to the above reasons.
However, in our study, the amount of increased CP content differed in the case of both microorganisms, showing that the type of fungal microorganisms may have affected the degree of increased CP. As overal the
P. ostreatus and P. chrysosporium fermentations were
found useful for increasing CP of tomato waste products.
The content of EE in tomato pomace was nearly consumed by the strain of P. ostreatus in our study. P.
ostreatus was also previously shown to reduce the EE
contents of oil-seed meal (Yasar and Tosun, 2018). The reduced lipid content could be due to the accumulation of lipids by some strains of fungal microorganisms which have lipase enzyme activity (Lateef et al., 2008; Agbo and Prah, 2014; Altop et al., 2019) since some fungal microorganisms should assimilate lipids from the fermenting substrates in order to produce other biomasses (Tinoco et al., 2011; Iandolo et al., 2011; Frexio et al., 2012; Jannathulla et al., 2018). In contrast P. chrysosporium significantly increased the EE content of tomato pomace. This increase in EE is thought to be due to the release of lipolytic enzymes
that break down glycerol and fatty acids in some microorganisms. Because Onweluza and Nwabugwu (2009) reported that the EE increases when millet
(Pennisetum americanum) and Pigeon pea (Cajanus
cajan) fermented and as a reason for the secreted
lipolytic enzymes during fermentation. However, in the previous studies, EE decreases; and lipolytic enzyme secretion and the increase in EE in fermentation are not sufficient in the literature. Some microorganisms generally use easily soluble carbohydrates such as starch and sugar to meet carbon requirements during fermentation, and then prefer to use complex carbohydrates or other nutrient as carbon sources (Papagianni, 2007; Altop et al., 2019). And as well Xie et al. (2016) and Altop et al. (2019) reported that during solid state fermentation, fungals release enzymes such as cellulase, hemicellulose, which break down structural carbohydrates, and these enzymes break down structural carbohydrates. It was concluded that the microorganism used as a carbon source as a reason for the decrease in RS and HS content in fermentation of tomato pomace with P. chrysosporium. On the other hand, P. chrysosporium is a white-rot-fungus, the release of enzymes that break down structural carbohydrates is thought to be high and therefore the HS content of tomato pomace may be reduced. In P. ostreatus fermentation, the HS content did not change, but the RS content decreased significantly, suggesting that this microorganism did not use either the carbon requirement of P.
chrysosporium or the HS as a carbon source or does not
release enough enzymes to break down structural carbohydrates
There was no significant effect on the contents of ANFs (tannin and pectin) of tomato pomace of the fermentation carried out without fungal microorganisms in this study (P>0.05) (Table 4). However, fermenting tomato pomace with P. ostreatus
and P. chrysosporium significantly influenced these
decrease in the tannin content at the 14 day of the fermentations carried out both fungal microorganisms (P<0.05). Fermentation of tomato pomace with P.
ostreatus did not significantly change the pectin
content (P>0.05). However, there was a significant sporadic effect of P. chrysosporium fermentation on the pectin content, which was increased (P<0.05) by 48% at 14 day and then decrease (P<0.05) by 28% at 21 day of in the pectin contents.
The pectin and tannin are usually available in the tomato peel tissue, and its degradation is very important to free up important biologically active
compound such as phenolic compounds and lycopene with antioxidant property (Lavecchia and Zuorro, 2008; Rodríguez-Fernández et al., 2011; Saleh et al., 2018). Biz et al., (2016) reported that they produce pectinase enzyme by fermenting agricultural by-products with Aspergillus oryzae. Enzyme production in the presence of pectin in the fermentation of microorganisms in the presence of pectin to break down the enzyme is connected to secrete. In our studythe pectin content of tomato peels was extracted and released into the matrix of fermentation substrate at the 14 day of fermentation with both fungal species, due to a possible increase in the activitiy of pectinase. Table 4. Influence of fungal fermentation on the contents of tannin and pectin of tomato pomace
Çizelge 4. Fungal fermantasyonunun domates posasının tannin ve pektin içeriğine etkisi Microorganisms Days Experiments I
(P. ostreatus) Experiments II(P. chrysosporium) Experiments (Control) III
Tannin, % 0 13.17±0.84a 13.17±0.84a 13.20±0.02 7 4.91±0.10b 6.55±0.34b 13.00±0.50 14 4.98±0.80b 5.52±1.43b 13.00±0.20 21 5.36±0.12b 6.99±0.36b 12.90±0.01 Pectin, % 0 3.43±0.31 3.43±0.28b 3.40±0.02 7 4.19±0.46 3.77±0.69b 3.32±0.06 14 3.91±0.19 5.09±0.49a 3.64±0.09 21 4.47±0.81 2.47±0.23c 3.40±0.05
a,b,c Different superscripts showed significant differences between the fermentation periods at each of the column parameters. Finally, the degradation of tomato peels could be said
to be completed by fungal fermentations. This mechanism was more pronounced with the fermentation using P. chrysosporium. Thus, it could possible that some important phenolic compounds are released during the fungal fermentations and this lead to a possible increase of antioxidant capacity.
The tannin content of tomato pomace decreased in fermentation with both fungal microorganisms. In previous studies, it has been reported that in fermentation microorganisms break down tannin into smaller molecules such as gallic acid, catechin, glucose and gallicatechin (Rodriguez et al., 2008; Nazarni et al., 2016; Shang et al., 2019). In this study, it is thought that the decrease in tannin content secretes tannase enzyme and the tannin content is converted to gallic acid, catechin, glucose or gallicatechin.
CONCLUSION
As a result, the fermentation of tomato pomace by fungal microorganism yielded an enrichment of nutritional qualities and added some biologically functional compounds. The fermented tomato pomace with improved nutritional qualities is holding a great potential in farm animal nutrition.
ACKNOWLEDGEMENT
We gratefully acknowledged the TUBITAK, Turkey for their financial support of the study (VHAG: 214O629 - 1001 project).
Statement of Conflict of Interest
Authors have declared no conflict of interest. Author’s Contributions
The contribution of the authors is equal. REFERENCES
Abdollahzadeh F, Pirmohammadi R, Fatehi F, Bernousi I 2010. Effect of feeding ensiled mixed tomato and apple pomace on performance of Holstein dairy cows. Slovak Journal of Animal Science, 43(1): 31-35.
Afşin M 2010. Katı faz fermantasyon (solid state fermantation; SSF) yöntemiyle Bacillus
licheniformis ATCC 14580'den proteaz üretimi.
Dicle Üniversitesi Fen Bilimleri Enstitüsü Biyoloji Anabilim Dalı, Yüksek Lisans Tezi 95 sy.
Agbo NW, Prah CD 2014. Evaluation of fermentation period on the proximate composition and tannin concentration of sheanut (Vitellaria paradoxa) meal. Journal of Microbiology and Biotechnology Research, 4(1): 21-27.
Aguilar CN, Aguilera-Carbo A, Robledo A, Ventura J, Belmares R, Martinez D, Rodriguez R, Contreras-Esquivel JC 2008. Production of antioxidant nutraceuticals by solid-state cultures of pomegranate (Punica granatum) peel and creosote bush (Larrea tridentata) leaves. Food Technology and Biotechnology, 46(2): 218–222.
Ajila CM, Brar SK, Verma M, Tyagi RD, Valéro JR 2011. Solid-state fermentation of apple pomace using Phanerocheate chrysosporium – Liberation and extraction of phenolic antioxidants. Food Chemistry, 126(3): 1071-1080.
Al-Betawi 2005. Preliminary study on tomato pomace as unusual feedstuffin broiler diets. Pakistan Journal of Nutrition, 4(1): 57-63.
Altop A, Coskun I, Filik G, Kucukgul A, Bekiroglu YG, Cayan H, Gungor E, Sahin A, Erener G 2018. Amino acid, mineral, condensed tannin, and other chemical contents of olive leaves (Olea europaea L.) processed via solid-state fermentation using selected Aspergillus niger strains. Ciencia e Investigacion Agraria, 45(2): 220-230.
Altop A, Gungor E, Erener G 2019. Improvement of Nutritional Quality of Some Oilseed Meals Through Solid-State Fermentation Using Aspergillus niger. Turkish Journal of Agriculture - Food Science and Technology, 7(9): 1411-1414.
AOAC 2005. Official Methods of Analysis. 18th ed., https://www.aoac.org/AOAC_Prod_Imis/AOAC_Me mber/Default.aspx?WebsiteKey=2e25ab5a-1f6d- 4d78-a498-19b9763d11b4&hkey=8fc2171a-6051-4e64-a928-5c47dfa25797.
Assi JA, King AJ 2008. Manganese amendment and Pleurotus ostreatus treatment to convert tomato pomace for inclusion in poultry feed, Poultry Science, 87(9):1889-1896.
Azza AM, El-Safy SF, Eman HFA 2013. Improvement of nutritional quality and antioxidant activities of fermented wastes by Saccharomyces cervisiae, Bacillus subtilis and Pleurotus salmoneo-stramineus. 3rd International Conference on Biotechnology and Its Applications in Botany and Microbiology, 17-18 April 2018, Cairo, Egypt. Bennamoun L, Hiligsmann S, Dakhmouche S,
Ait-Kaki A, Labbani FK, Nouadri T, Meraihi Z, Turchetti B, Buzzini P, Thonart P 2016. Production and properties of a thermostable, ph stable exo-polygalacturonase using Aureobasidium pullulans ısolated from Saharan Soil of Algeria grown on tomato pomace. Foods, 5(4): E72.
Biz, A, Finkler ATJ, Pitol LO, Medina BS, Krieger N, Mitchell DA 2016. Production of pectinases by solid-state fermentation of a mixture of citrus waste and sugarcane bagassein a pilot-scale packed-bed bioreactor. Biochemical Engineering Journal, 111(8): 54-62.
Chemesova II, Chizhikov DV 2004. Determination of tannin content in rhizomes of Comarum palustre L. and its tincture by spectrophotometric method. Rastitel'nye Resursy, 40: 122-130.
Dei HK, Rose SP, Mackenzie AM, Amarowicz R 2008. Growth performance of broiler chickens fed diets containing shea nut (Vitellaria paradoxa, Gaertn.) meal fermented with Aspergillus niger. Poultry Science, 87(9): 1773–1778.
Díaz-Godínez G 2012. Production of laccases by Pleurotus ostreatus in solid-state and submerged fermentation. Biotechnology of Microbial Enzymes, Nova Science Publishers, A: 99-121.
Ergun SO, Urek RO 2017. Production of ligninolytic enzymes by solid state fermentation using Pleurotus ostreatus. Annals of Agrarian Science, 15(2): 273-277.
FAO, 2016. Food and Agrıculture Organization of the United Nations. FAO Statics Data 2016, http://www.fao.org/faostat/en/#data.
Freixo MR, Karmali A, Arteiro JM 2008. Production and chromatographic behavior of polygalacturonase from Pleurotus ostreatus on immobilized metal chelates. Process Biochemistry,43(5):531-539. Freixo MR, Karmali A, Arteiro JM 2012. Production,
purification and characterization of laccase from Pleurotus ostreatus grown on tomato pomace. World Journal of Microbiology & Biotechnology, 28(1): 245–254.
Gassara F, Brar SK, Tyagi RD, Verma M, Surampalli RY 2010. Screening of agro-industrial wastes to produce ligninolytic enzymes by Phanerochaete chrysosporium. Biochemical Engineering Journal,49(3):388-394.
Goyal M, Soni G 2011. Production and characterization of cellulolytic enzymes by Pleurotus florida. Mycosphere, 2(3): 249–254.
Iandolo D, Piscitelli A, Sannia G, Faraco V 2011. Enzyme production by solid substrate fermentation of Pleurotus ostreatus and Trametes versicolor on tomato pomace. Applied Biochemistry and Biotechnology, 163(1): 40-51.
Jannathulla R, Dayal JS, Ambasankar K, Muralidhar M. 2018. Effect of Aspergillus niger fermented soybean meal and sunflower oil cake on growth, carcass composition and haemolymph indices in Penaeus vannamei Boone, 1931. Aquaculture, 486(3): 1-8.
King AJ, Zeidler G 2004. Tomato pomace may be a good source of vitamin E in broiler diets. California Agrıculture, 58(1):59-62.
Knoblich M, Anderson B, Latshaw D 2005. Analyses of tomato peel and seed by products and their use as a source of carotenoids. Journal of the Science of Food and Agriculture, 85(7): 1166–1170.
Kurt S, Buyukalaca S 2010. Yield performances and changes in enzyme activities of Pleurotus spp. (P. ostreatus and P. sajor-caju) cultivated on different agricultural wastes, Bioresource Technology, 101(9): 3164-3169.
Lateef A, Oloke JK, Kana EBG, Oyeniyi SO, Onifade OR, Oyeleye AO, Oladosu OC, Oyelami AO 2008. Improving the quality of agro-wastes by solid-state fermentation: enhanced antioxidant activities and nutritional qualities. World Journal of Microbiology and Biotechnology, 24(10): 2369–2374.
Lavecchia R, Zuorro A 2008. Improved lycopene extraction from tomato peels using cell-wall degrading enzymes. European Food Research and Technology, 228(2): 153-158.
Liu J, Cai Y, Liao X, Huang Q, Hao Z, Hu M, Zhang D, Li Z 2013. Efficiency of laccase roduction in a 65-L air-lift reactor for potential green industrial and environmental application. Journal of Cleaner Production, 39(2): 154-160.
Lu Z, Wang J, Gao R, Ye F, Zhao G 2019. Sustainable valorisation of tomato pomace: A comprehensive review. Trends in Food Science & Technology, 86(4):172-186.
Mirzaei-Aghsaghali A, Maheri-Sis N 2008. Nutritive value of some agro-industrial by-products for ruminants. World Journal of Zoology, 3(2): 40-46. Mukherjee R, Chakraborty R, Dutta A 2016. Role of
fermentation in improving nutritional quality of soybean meal a review. Asian-Australasian Journal of Animal Sciences, 29(11): 1523-1529.
Nazarni R, Purnama D, Umar S, Eni H 2016. The effect of fermentation on total phenolic, flavonoid and tannin content and its relation to antibacterial activity in jaruk tigarun (Crataeva nurvala, Buch HAM). International Food Research Journal 23(1): 309-315
Oboh G, Akindahunsi A 2003. Biochemical changes in cassava products (flour & gari) subjected to Saccharomyces cerevisiae solid media fermentation. Food Chemistry, 82(4): 599–602. Okpako C, Ntui V, Osuagwu A, Obasi F 2008.
Proximate composition and cyanide content of cassava peels fermented with Aspergillus niger and Lactobacillus rhamnosus. Journal of Food Agriculture and Environment, 6(2): 251-255. Özşölen F 2010. Production of ligninolytic enzymes
with solid state fermentation. Eskişehir Osmangazi Üniversitesi Fen Bilimleri Enstitüsü Biyoloji Anabilim Dalı, Yüksek Lisans Tezi, 85 sy.
Papagianni M 2007. Advances in citric acid fermentation by Aspergillus niger: biochemical aspects, membrane transport and modeling. Biotechnology Advances, 25(3):244-63.
pomegranate pomace extract
Raimbault M 1998. General and microbiological aspects of solid substrate fermentation. Electronic Journal of Biotechnology, 1(3): 717-732.
Rashad MM, Abdou HM, Mahmoud AE, Nooman MU 2009. Nutritional analysis and enzyme activities of Pleurotus ostreatus cultivated on citrus limonium and carica papaya wastes. Australian Journal of Basic and Applied Sciences, 3(4): 3352-3360. Rashad MM, Abdou HM, Shousha WGH, Ali MM,
El-Sayed NN 2010. Purification and characterization of extracellular polygalacturonase from Pleurotus ostreatus using citrus limonium waste. Journal of Applied Science Research, 6(1): 81-88.
Ravichandran S, Vimala R 2012. Solid state and
submerged fermentation for the production of bioactive substances: a comparative study. International Journal of Science and Nature, 3(3):480-486.
Rodríguez H, Rivas B, Cordovés CG, Muñoz R 2008. Degradation of tannic acid by cell-free extracts of Lactobacillus plantarum. Food Chemistry 107(2): 664–670.
Roja HN, Munishamanna KB, Veena R, Palanimuthu V 2017. Solid state fermentation of tomato pomace waste by different lactic acid bacteria and yeast strains for quality and nutritional improvement. Agrıculture Update, 12(2):347-352.
Saleh H, Golian A, Kermanshahi H, Mirakzehi MT 2018. Antioxidant status and thigh meat quality of broiler chickens fed diet. Italian Journal of Animal Science, 17(2): 386–395.
Shanga YF, Cao H, Ma Yl, Zhang C, Ma F, Wang CX, Ni XL, Lee WJ, Wei ZJ 2019. Effect of lactic acid bacteria fermentation on tannins removal in Xuan Mugua fruits. Food Chemistry, 274(5): 118-122. Singhaniaa RR, Patel AK, Soccol CR, Pandey A 2009.
Recent advances in solid state fermentation. Biochemical Engineering Journal, 44(1): 13–18. SPSS 2013. IBM SPSS Statistics 22.0 for Windows.
Armonk, NY.
Tinoco R, Acevedo A, Galindo E, Serrano-Carreo’n L (2011). Increasing Pleurotus ostreatus laccase production by culture medium optimization and copper/lignin synergistic induction. Journal of Industrial Microbiology and Biotechnology, 38(4): 531-540.
TS ISO 21527-2: 2008. Microbiology of food and animal feeding stuffs — Horizontal method for the enumeration of yeasts and moulds — Part 2: Colony count technique in products with water activity less than or equal to 0,95; TS ISO 21527-2: 2008, Avaliable from: https://www.iso.org/standard/ 38276.html.
Ülger I, Kaliber M, Ayaşan T, Küçük O 2018. Chemical composition, organic matter digestibility and energy content of apple pomace silage and its combination with corn plant, sugar beet pulp and pumpkin pulp*. South African Society for Animal Science, 48(3): 497-503.
Verma P, Madamwar D 2002. Production of ligninolytic enzymes for dye decolorization by cocultivation of white-rot fungi Pleurotus ostreatus and Phanerochaete chrysosporium under solid-state fermentation. Applied Biochemistry and Biotechnology, 102(1-6): 109-118.
Villas-Boas SG, Esposito E, De Mendonca MM 2003. Bioconversion of apple pomace into a nutritionally enriched substrate by Candida utilis and Pleurotus ostreatus. World Journal of Microbiology and Biotechnology, 19(5):461-467.
Wadhwa M, Bakshi MPS 2016. Application of Waste-Derived Proteins in the Animal Feed Industry. (In:
Gurpreet Dhillon, editors: Protein Byproducts, Oxford: Academic Press; 2016) 161-192.
Waldbauer K, McKinnon R, Kopp B 2017. Apple Pomace as Potential Source of Natural Active Compounds. Planta Medica, 83(12-13):994-1010. Wang XP, Zhang WY 1999. Study on the determination
and utilization of the pectin in haw. Quarterly of Forest By-Product and Speciality in China, 9(3): 13-14.
Weiss WP, Frobose DL, Koch ME 1997. Wet tomato pomace ensiled with corn plants for dairy cows. Journal of Dairy Science, 80(11): 2896–2900. Xie PJ, Huang LX, Zhang CH, Zhang YL 2016.
Nutrient assessment of olive leaf residues processed by solid-state fermentation as an innovative feedstuff additive. Journal of Applied Microbiology, 121(1):28-40.
Yasar S, Tosun R 2018. Nutritional, enzymatic and antioxidant enrichment of apple pomace by fungal fermentations. 10. International Animal Science Conference, 25-27 October 2018, Antalya.
Yasar S, Tosun R 2019. Increasing the nutritional qualities of tomato pomace by yeast fermentation. 4th International conference on advances in natural & applied sciences June 19-22, 2019, Ağrı.
Yoon LW, Ang TN, Ngoh GC, Chua ASM 2014. Fungal solid-state fermentation and various methods of enhancement in cellulase production. Biomass and Bioenergy, 67(5):319-338.
Ziaei N, Molaei S 2010. Evaluation of nutrient digestibility of wet tomato pomace ensiled with wheat straw compared to alfaalfa hay in kermani sheep. Journal of Animal and Veterinary Advances, 9(4): 771-773.