T.C.
EGE ÜNİVERSİTESİ TIP FAKÜLTESİ
ÇOCUK SAĞLIĞI VE HASTALIKLARI ANA BİLİM DALI PROF. DR. SAVAŞ KANSOY
OSTEOKONDRODİSPLAZİLERDE SERUM C-TİP
NATRİÜRETİK PEPTİD PLAZMA DÜZEYİ
UZMANLIK TEZİ Dr. Sirmen KIZILCAN ÇETİN
DANIŞMAN
Prof. Dr. Şükran DARCAN
İZMİR 2018
ÖNSÖZ VE TEŞEKKÜR
Doğduğum, büyüdüğüm, dağlarında çiçek açan şehrin güzel hastanesi... Daha çocukken düştü bu sevda minik kalbime, büyüyecektim ve doktor olacaktım. İlk defa çocukken hastası olarak girdim kapısından. Sonrasında o büyük gün gelmişti; Ege Üniversitesi Tıp Fakültesi’nin kapısından öğrencisi olarak girecektim. Sevinçten ağlamak ne demek o gün anlamıştım.
Ferdi olmaktan büyük onur duyduğum Ege Pediatri Ailesi’nin temellerini atan rahmetli Sabiha Cura Özgür Hocamız başta olmak üzere, aileye girdiğim andan itibaren beni kucaklayan, sevdalısı olduğum bu mesleği öğrenmem için büyük uğraş veren, geldiğim bu noktada, başardıklarım ve başaracaklarım üzerinde pahabiçilemez emekleri olan, her zaman minnet ve şükran ile anacağım saygıdeğer hocalarıma ve Anabilim Dalı Başkanımız Prof. Dr Savaş Kansoy’a.
Daha öğrenciyken nasıl anamnez alınır, hastaya nasıl dokulunur, nasıl muayene edilir, iyi hekim nasıl olunuru bana öğreten; öğrenciyken kliniğe adım attığımda da, diplomamı alırken de, asistan olup kliniğe dönüşümde de hep yanımda olan, kapısını çaldıgımda hep vakit yaratan, bir sorduğumda bin öğreten, nöbet çıkışı görüp çocuğum biraz soluklan açken vizit verme diyen, vizyonumu geliştirmek için yurtdışına gitmek istediğimi duyduğundaki mutluluğu aklımdan çıkmayan ve yurtdışında bile bir anımda yalnız hissettirmeyen, endokrinolojiyi onunla daha çok sevdiğim, benim için idol olan ve “Ben yetiştirdiğim öğrencime bildiğimi tamamen aktarmalıyım ve onun bunu geliştirmesini desteklemeliyim. Onun benden bir adım öteye geçtiğini gördüğümde o zaman başarılı bir hoca olmuşumdur.” sözünün sahibi, bu tezdeki emeği ve desteğinden öte, üzerimde pahabiçilemez emeği olan çok saygıdeğer hocam Prof. Dr. Şükran Darcan’a.
Bitmek tükenmek bilmeyen enerjisiyle her zaman araştıran, her zaman sorgulayan ve sorgulayan beyinlerin yetişmesi için çabalayan, diyabet denildiğinde akıllara gelen ilk isim olan, her ne öğrenmek istersem fazlasını aktaran, dünyanın her neresinde olursam olayım desteğini hep hissettiğim, daha iyi bir hekim olmak, vizyonumu geliştirmek adına bana her türlü fırsatı tanıyan, gerek tezimizin hazırlanmasında gerekse öğrencilik ve asistanlık sürecinde eğitim öğrenimimde sonsuz katkıları olan çok değerli hocam Prof. Dr. Damla Gökşen’e.
Hep dinamik hep güleryüzlü olan, vakalar üzerine tartışmaktan keyif aldığım, en karmaşık gelen konuyu bile oldukça basit ve anlaşılabilir anlatan, her an müsait ve her zaman destek
olan, daha fazlasını öğretmek için elindeki her şeyi paylaşan, endokrin sevgisine genetiği de katan sevgili hocam Doç. Dr. Samim Özen’e.
Siyah ve beyazdan oluşan filmlere can katan, kapısı her zaman sonuna kadar açık olan, çocukları radyasyondan korumak adına “Primum non nocere”yi iliklerimize işleten, katıldığı konseylerde son noktayı koyarken herkesi kendisine hayran bırakan, gecenin 04:00’ünde bile hastaneye uğrayıp hatrımızı soran, bir sorduğumuzda bin öğreten, tezimizin hazırlanma süresince her bir hastanın grafisini büyük itinayla irdeleyen, saygıdeğer hocam ve BALdaş abim olan Prof. Dr. Hüdaver Alper’e.
Her zaman güleryüzüyle, desteğiyle, çalışkanlığıyla bana örnek olan, içi genetik sevgisiyle dolu olan ve beni de genetik sevdalısı yapan, hayata OMIM, Varsome gözüyle bakmamı sağlayan ve gerçekten birşeyler üretmek isitiyorsam çok iyi bir genetik bilgisine sahip olmam gerektiğini bana öğreten canım ablam Uzm. Dr. Esra Işık’a.
Çalışma verilerimizin değerlendirilmesinde yardımları olan Ege Üniversitesi Tıp Fakültesi Biyoistatistik ve Tıbbı Bilişim Anabilim Dalı öğretim üyeleri ve asistanlarına.
Katılımcıları karşılamamızda ve dosyalarını taramamızda yardımları olan Endokrinoloji BD sekreterleri Aylin Kabak, Zekeriya Bey ve Serkan Bey’e, katılımcıların kanlarını canlarını acıtmadan, itinayla alan Perihan Hemşire Hanım ve Günay Hemşire Hanım’a, katılımcıların tetkiklerinin çalışılmasında emeği olan Endokrinoloji laborantı Emine Hanım’a ve çalışmaya katılan çocuklarımız ve ailelerine, sonsuz teşekkürlerimi sunarım.
Dört yıl boyunca gecelerini gündüzlerine katan, nöbetlerde bir telefon ucunda olan, eğitimime pahabiçilemez katkıları olan uzman abi ve ablalarıma; stajer ve intörnleri olarak çalışmaktan keyif aldığım ve Ege Pediatri ailesinin sıcaklığını bana hissettiren, asistanlığa başladığımda bana “sevgi pıtırcığı” lakabını takan kıdemlilerime; çalışmaktan keyif aldığım ve hep desteklerini, ekip ruhunu hissettiğim eş kıdem ve alt kıdem çalışma arkadaşlarıma; gece gündüz ekip olarak çalışmanın keyfini yaşatan çok değerli hemşire arkadaşlarıma; güleryüzlü bilim dalı sekreterlermize ve işlerin yolunda gitmesi için çabalayan Ege Pediatri Ailesi tüm personeline teşekkürü borç bilirim.
Gülüşleri ile her türlü yorgunluğumu alan, benimle oyun oynayan, resimler çizen, boynuma sarılan, çok sevdiğim, her biri ayrı melek olan çocuk hastalarımıza ve çocukken hasta olduğumda beni iyileştiren, tüm şevkat ve ilgileriyle beni bu mesleği çocuk yaşta sevdiren çocuk doktorlarıma çok teşekkür ederim.
Bugünlere gelmemde paha biçilemez emeği olan, bana güvenen, inanan, her zaman destek olan, sevincime benden çok sevinen, üzüntüme benden çok üzülen, maddi-manevi hep yanımda olan, var olma sebebim canım annem Nevin, babam Eşref ve kardeşim Yiğit Kızılcan’a; 12 yıldır hayat arkadaşım olan, ÖSS’ye hazırlık sürecini birlikte göğüslediğimiz, tıp fakültesi sıralarında da, asistanlık eğitim sürecimde de bana hep destek ve anlayış gösteren, Ege Pediatri’de eğitimimi almak uğruna yıllarca ayrı yaşadığımız, zekası ve yaptığı
işlerle kendisine hayran kaldığım, her başarımda katkısı olan, her zaman bana inanan, güvenen ve destek olan biricik arkadaşım, sırdaşım, dostum, can yoldaşım, eşim Saygın Çetin’e sonsuz teşekkür ederim.
Bu çalışmayı destekleyen Ege Üniversitesi Bilimsel Araştırma Projeleri Koordinatörlüğü’ne teşekkür ederim.
Gönül hiç ayrılmak istemese de, geri dönüp doyasıya eğitim almaya devam etmeyi arzu etsem de, olur da dönememek varsa gelecekte; Egeli olmanın onuruyla, Egeli olmaya yakışır şekilde, dünyanın neresinde olursam olayım, minik kalpler için ilk günkü heyecanımla çalışacağımın bilinmesini isterim. Boşa söylenmemiş bir sözmüş; “Kalbim Ege’de kaldı...” Saygı ve sevgilerimle.
Dr Sirmen Kızılcan Çetin Temmuz 2018, İzmir
ÖZET
İskelet displazisi; anormal enkondral osifikasyon ile giden, heterojen bir hastalık grubu olup tiplendirmesi oldukça karmaşıktır. Natriüretik peptid ailesinden, C-tip natriüretik peptid (CNP)’nin kemik gelişiminde rol oynadığı gösterilmiştir. İskelet displazi tiplerinden bazılarında CNP düzeyleri yüksek bulunmuştur. Bu çalışmada amaç, kemik gelişiminde etkin olan CNP’nin iskelet displazi tipleri için belirteç olarak kullanılabilme olasılığını değerlendirmek, tiplendirmesini sağlamada rolünü araştırmaktır. Yaşları 0,5-18 yaş olan 75 iskelet displazili hastadan katılmayı kabul eden 37 hasta çalışmaya alındı. Olguların öyküleri alınarak,fizik bakı ve antropometrik ölçümleri yapıldı, kemik surveyleri değerlendirildi. Yaş grupları benzer, 49 sağlıklı çocuk kontrol grubunu oluşturuldu. Hasta ve kontrol grubunda plazmada CNP (ng/ml) ELISA yöntemi ile çalışıldı. Hasta grubun 11’i kız (%29,7), 26’sı erkek (%70,3); sağlıklı grubun 24’ü kız (%48,9), 25’i erkek (%51,1) idi. Hasta grubu; akondroplazili 17 (% 45,9), spondiloepifiziyal displazili 6 (%16,2), metafizyel displazili 5 (%13,5), epifizyel displazili 2 (%5,4), hipokondroplazili 2 (%5,4), akromezomelik displazili 1 (%2,7) ve sınıflandırılması yapılamayan 4 (%10,8) olgudan oluşmakta idi. Sonuçlar; hasta grubunun boy SDS değeri; -4,58±2,87 (n=37), sağlıklı grupta 0,05±0,79 (n=49) saptanmıştır (p<0,05). Hastalarda CNP konsantrasyonu ortanca değeri ile sağlıklı grubun ortanca değeri benzer bulundu (p=0,20). Sağlıklı grup (n=49) ile hasta grupta yer alan akondroplazi hastalarının (n=17) CNP konsantrasyon ortanca değeri, sağlıklı gruba oranla daha yüksek idi (p=0,032). Tüm katılımcılarda (erkek=51; kız=35 ) cinsiyetler arasında CNP konsantrasyonu ortancaları arasında anlamlı bir fark saptanmadı (p=0,082). Hasta grupta klinik tanılara göre CNP değeri akondroplazili hasta grubunda yüksek olmasına karşın; diğer iskelet displazili hastalarının sayısı yetersiz olduğundan, o grupta istatistiksel değerlendirme yapılamadı. Sonuç: CNP tüm iskelet displazilerinin tanı ve tiplendirmesinde kullanılabilir bir belirteçtir demek için; daha çok sayıda olgunun olduğu ve diğer moleküler genetik analizlerinin de yapıldığı çalışmalara gereksinim vardır.
ABSTRACT
Skeletal dysplasia is a heterogeneous group of disease, leading to abnormal enchondral ossification and typing is quite complex. C-type natriuretic peptide (CNP), one of the members of the natriuretic peptide family, has been implicated to play a role in bone development. CNP levels were high in some types of the skeletal dysplasia. The aim of this study is to evaluate the possibility of using CNP, which is effective in bone development, as a marker for skeletal dysplasia types and to investigate its role in typing. Participants were thirty-seven patients [ages 6 months to 18 years (26 girls, 11 boys)], who accepted to participate in the study from 75 skeletal dysplasia patients. All subjects were physically examined and anthropometric measurements were obtained, bone surveys were evaluated. A control group of 49 healthy children [ages 6 months to 18 years (24 girls, 25 boys)], was included. ELISA method was used to assess CNP (ng/ml) plasma levels. The patient group consisted of 17 patients (45,9%) with achondroplasia, 6 patients (16,2%) with spondyloepiphysial dysplasia , 5 patients (13,5%) with metaphysical dysplasia, 2 patients (5,4%) with epiphysial dysplasia, 2 patients (5,4%) with hypochondroplasia, one patient with acromesomelic dysplasia (2,7%) and 4 patients (10,8%) with unclassified skeletal dysplasia. Results: The SD scores (SDS) of the patient group was -4,58 ± 2,87 (n=37) and 0,05 ± 0,79 (n=49) in the healthy group (p<0,05). The median level of CNP concentration of patients was similar to the median level of healthy group (p=0,20). CNP concentration median value of healthy group (n=49) and achondroplasia patients (n=17) was higher than healthy group (p=0,032). There was no significant difference between CNP concentration medians of all the participants (male=51; female=35) (p=0,082). Although CNP value was higher in patients with achondroplasia according to clinical diagnosis in the patient group, as the number of other skeletal dysplasia patients was insufficient, statistical evaluation was not possible. Conclusion: Patient with achondroplasia has elevated plasma levels of CNP. In order to use CNP as a marker for diagnosing and typing all skeletal dysplasia, more clinical studies with molecular genetic analyzes are needed.
İÇİNDEKİLER
ÖNSÖZ VE TEŞEKKÜR ... ii
ÖZET ... v
ABSTRACT ... vi
TABLO LİSTESİ ... xii
ŞEKİL LİSTESİ ... xiv
KISALTMALAR ... xvi 1 GİRİŞ VE AMAÇ ... 1 2 GENEL BİLGİLER ... 3 2.1 Natriüretik peptidler ... 3 2.1.1 ANP ... 4 2.1.2 BNP ... 5 2.1.3 CNP ... 5 2.1.4 DNP ... 6 2.1.5 Osteokrin/ Musklin ... 6
2.2 Natriüretik Peptid Reseptörleri ... 7
2.3 Büyüme Üzerine Etkili Diğer Hormonlar ... 12
2.4 İskelet Displazileri (OSTEOKONDRODİSPLAZİLER) ... 13
2.4.1 Giriş ... 13
2.4.2 Normal Kemik Gelişimi ... 14
2.4.3 İskelet Displazileri Sınıflandırılması ... 14
2.4.4 Kemikte Büyüme Bozukluğu İle Karakterize Olan İskelet Displazileri ... 18
2.4.4.1.1 Akondrogenezi I (Parenti-Fraccaro) Ve Akondrogenezi Iı (Langer-Saldino) ... 18 2.4.4.2 Akondroplazi ... 19 2.4.4.3 Hipokondroplazi ... 22 2.4.4.4 Punktat Kondrodisplazi ... 22 2.4.4.5 Diastrofik Displazi ... 23 2.4.4.6 Metatropik Displazi ... 24 2.4.4.7 Mezomelik Displazi ... 24 2.4.4.8 Akromezomelik Displazi ... 25
2.4.4.9 Kongenital Spondiloepifizer Displazi ... 25
2.4.4.10 Kniest Sendromu ... 26
2.4.4.11 Kondroektodermal Displazi ‘’Ellisvan Creveld Sendromu’’ ... 26
2.4.4.12 Asfiksi Yapan Torasik Displazi (Jeune Sendromu) ... 26
2.4.4.13 Kleidokranial Displazi ... 27
2.4.4.14 Spondilo-Epifizo-Metafizer Displaziler ... 27
2.4.4.15 Akrodisplaziler ... 29
2.4.4.16 Jeneralize Platispondililer ... 30
2.4.4.17 Spondiloepifizyel Displazi ... 30
2.4.5 Kıkırdak Veya Fibröz Doku Büyümesinde Düzensizlik Gösteren Osteokondrodisplaziler ... 31
2.4.5.1 Hemimelik Epifizer Displazi ... 31
2.4.5.2 Çoklu Kıkırdak Ekzostozu ... 31
2.4.5.3 Diskondroplazi (Enkondromatoz, Çoklu İskelet Kondromatozis) ... 32
2.4.5.4 Maffucci Sendromu (Enkondromatoz ile Hemanjiomatoz) ... 32
2.4.5.5 Fibröz Displazi ... 32
2.4.5.6 Çerubizm ... 32
2.4.6 Dansite Değişikliği Veya Kemiğin Şekillenmesinde Bozukluk İle Karakterize Osteokondrodisplaziler ... 33
2.4.6.1 Osteogenesis İmperfekta ... 33
2.4.6.2 Osteopetrozis ... 34
2.4.6.4 Meloreostoz ... 35
2.4.6.5 Osteopoikiloz ... 35
2.4.6.6 Diafizer Displazi (Camurati-Engelmann Hastalığı) ... 36
2.4.6.7 Hiperfosfataz İle Osteoektazi ... 36
2.4.6.8 Osteodisplasti (Melnick-Needles Sendromu) ... 37
2.4.6.9 Genelleştirilmiş Kortikal Hipostoz (Endosteal Hiperostoz) ... 37
2.4.6.10 Psödoakondroplazi ... 37
2.4.6.11 Multiple Epifizyal Displazi ... 38
2.4.6.12 Jansen Metafizyel Kondrodisplazi ... 39
2.4.6.13 Blomstard Kondrodisplazi ... 39
2.4.6.14 Pyle Hastalıkları (Metafizeal Osteodisplazi) ... 39
2.4.6.15 Kraniyometafizal Displazi ... 39 2.4.6.16 Göz-Diş-Kemik Displazisi ... 40 2.4.7 Disostozlar ... 40 2.4.7.1 Kranial Disostoz ... 40 2.4.7.2 İzole Kraniosinostoz ... 41 2.4.7.3 Kombine Kraniosinostozlar ... 41
2.4.7.3.1 Kraniofasial Dizostoz (Crouzon Sendromu) ... 41
2.4.7.3.2 Akrosefalosindaktili ... 42
2.4.7.3.3 Yonca Yaprağı Kafatası (Clover Leaf Skull) ... 42
2.4.7.4 Fasial Dizostozlar ... 42
2.4.7.5 Mandibula-Fasial Dizostozlar ... 42
2.4.7.6 Mandibuler Hipoplazi (Pierre-Robin Sendromu) ... 43
2.4.7.7 Okulo-Mandibula-Fasial Sendrom (Hallermann-Streiff-François Sendromu) ... 43
2.4.7.8 Bazal Hücre Karsinom Sendromu ... 43
2.4.7.9 Spinal Dizostozlar ... 43
2.4.7.10 Klippel-Feil Sendromu ... 43
2.4.7.11 Spondilokostal Dizostoz ... 44
2.4.7.12 Omuz Ve Kalça Tultulması Olan Disostozlar ... 44
2.4.7.13 Osteoonikodizostoz (Tırnak-Patella Sendromu) ... 44
2.4.7.14 Ekstremiteleri Tutan Disostozlar ... 44
2.4.7.14.2 Konjenital Uzun Kemik Eğrilikleri ... 45
2.4.7.14.3 Kampomelik Sendrom ... 45
2.4.7.14.4 İzole Tibia Eğriliği ... 45
2.4.7.14.5 Ekstremitelerin Segment Anomalileri ... 45
2.4.7.14.6 Karpal Ve Tarsal Kemik Anomalileri ... 46
2.4.7.14.7 Metakarpal veya Metatarsal Falangeal Anomaliler ... 46 2.4.7.14.7.1 Brakidaktililer ... 46 2.4.7.14.7.2 Sinfalanjizm ... 46 2.4.7.14.7.3 Multiple Sinostoz ... 46 2.4.7.14.7.4 Polidaktili ... 46 2.4.7.14.7.5 Sindaktili ... 47 2.4.7.14.7.6 Kamptodaktili ... 47 2.4.7.14.7.7 Başparmak Anomalileri ... 47 2.4.7.14.7.8 Akroosteoliz ... 47
2.4.8 İskelet Displazilerinin Tanısı ... 47
2.4.8.1 Anamnez ... 48 2.4.8.2 Fizik Muayene ... 48 2.4.8.3 Biyokimyasal Çalışmalar ... 50 2.4.8.4 İskelet Radrografileri ... 51 2.4.8.5 Histopatolojik Değerlendirme ... 54 2.4.8.6 Prenatal Tanı ... 56 2.4.8.7 Genetik ... 57 2.4.9 Tedavi ... 59
2.4.9.1 Seçilmiş Displazilerin Farmakolojik Yönetimi ... 59
2.4.9.2 Büyüme Hormonu ... 59
2.4.9.2.1 Büyüme Hormonu Tedavi Dozu- Akondroplazi ... 60
2.4.9.2.2 Büyüme Hormonu Tedavi Dozu- Hipokondroplazi ... 61
2.4.9.2.3 Büyüme Hormonu Nörolojik Komplikasyon Arası İlişki ... 61
2.4.9.3 Bifosfonatlar ... 62
2.4.9.5 Enzim Replasman Tedavisi (ERT) ... 67
2.4.9.6 Cerrahi İşlemler ... 67
2.4.9.7 Psikolojik Destek Grupları ... 68
2.4.10 İskelet Displazilerine eşlik eden klinik ve labaratuvar bulgular ... 69
2.4.10.1 Hiperglisemi ... 69
2.4.10.2 Hipokalsemi ... 69
2.4.10.3 Hiperkalsemi ... 69
2.4.10.4 Cinsiyet Farklılaşma Bozukluğu ... 70
2.4.10.5 Adrenal Yetmezlik ... 70
3 GEREÇ VE YÖNTEM ... 71
3.1 Etik Kurul Onayı ... 71
3.2 Çalışma Grubunun Oluşturulması ve Değerlendirilmesi ... 71
3.2.1 Çalışmaya Dahil Edilme Kriterleri ... 72
3.2.2 Çalışmaya Alınmama Kriterleri ... 72
3.2.3 Çalışmadan Çıkarılma Kriterleri ve Bu Durumda Yapılacak Uygulamalar ... 73
3.3 Hasta Değerlendirmesi ... 73
3.4 İnsan CNP (2', 3'-siklik nükleotid 3'-fosfodiesteraz) ELISA Testi ... 73
3.4.1 Deney İlkeleri ... 74
3.4.2 Önlemler ... 74
3.4.3 Manuel Yıkama ... 75
3.4.4 Otomatik Yıkama ... 75
3.4.5 Örnek Toplama ve Depolama ... 75
3.4.6 Numune Seyreltme ... 76
3.4.7 Reaktif Hazırlama ve Depolama ... 76
3.4.7.1 Yıkama Tamponu ... 77
3.4.7.2 Standart ... 77
3.4.7.3 Biyotinle Etiketlenmiş Antikor Çalıştırma Çözeltisinin Hazırlanması ... 77
3.4.7.4 HRP-Streptavidin Konjugatı’nın (SABC) Hazırlanması ... 78
3.4.8 Deney İşlemleri ... 78
3.4.9 Özet... 79
4 BULGULAR ... 81 4.1 Demografik veriler ... 81 5 TARTIŞMA ... 96 6 SONUÇ VE ÖNERİLER ... 112 KAYNAKLAR ... 115 EKLER ... 121
Ek.1 Genetik iskelet sistemi hastalıklarının biyokimyasal, gentik ve radyolojik kriterlere göre sınıflaması ... 121
Ek.2 Etik Kurul Onayı ... 154
Ek.3 Bilgilendirilmiş Gönüllü Olur Formu – Hasta Grubu Ebeveyn(Form 17)... 159
Ek.4 Bilgilendirilmiş Kontrol Olur Formu (Form 17)-Kontrol Grubu Ebeveyn ... 164
Ek.5 Bilgilendirilmiş Hasta Olur Formu Çocukluk Yaş Grubu ( 6-12 Yaş Arası) –Hasta Grubu... 170
Ek.6 Bilgilendirilmiş Gönüllü Olur Formu Çocukluk Yaş Grubu ( 6-12 Yaş Arası)-Kontrol Grubu ... 174
Ek.7 Bilgilendirilmiş Gönüllü Olur Formu -Hasta Grubu Ergenler İçin ... 178
Ek.8 Bilgilendirilmiş Olur Formu-Kontrol Grubu (Ergenler İçin 13-18 Yaş) ... 182
Ek.9 Olgu İzleme Formu ... 186
TABLO LİSTESİ
Tablo 1. Hasta ve sağlıklı çocuklarda antropometrik ölçümlerin diğer
gruplarla karşılaştırılması ... 83
Tablo 2. Akondroplazili ve sağlıklı çocuklarda antropometrik ölçümlerin
diğer gruplarla karşılaştırılması ... 84
Tablo 3. Hasta ve sağlıklı çocuklarda CNP konsantrasyonu
karşılaştırılması ... 85
Tablo 4. Akondroplazili ve sağlıklı çocuklarda CNP konsantrasyonu
karşılaştırılması ... 85
Tablo 5. Cinsiyete bağlı CNP konsantrasyon (ng/ml) değerlerinin
karşılaştırılması ... 86
Tablo 6. Cinsiyete bağlı CNP konsantrasyon (ng/ml) değerlerinin
karşılaştırılması (Hasta - Sağlıklı kıyaslamalı) ... 88
Tablo 7. CNP konsantrasyonu; hasta ve sağlık gruplarda, yaşlar arasındaki
değişimi ... 88
Tablo 8. CNP konsantrasyonu; akondroplazili hasta ve sağlık gruplarda,
yaşlar arasındaki değişimi ... 89
Tablo 9. Kruskal-Wallis Test İstatistiği ... 90
Tablo 10. İskelet Displazilerinde CNP Konstantrasyon Karşılaştırılması ... 90
Tablo 11. Hasta ve Sağlıklı Antropometrik Ölçümleri ile CNP
Konsantrasyon Korelasyon ... 91
Tablo 12. Hasta Biyokimyasal Parametreleri ile CNP Konsantrasyon
Korelasyonu ... 92
Tablo 13. Akondroplazili ve Sağlıklı Antropometrik Ölçümleri ile CNP
Konsantrasyon Korelasyonu ... 93
Tablo 14. Akondroplazi Biyokimyasal Parametreleri ile CNP
ŞEKİL LİSTESİ
Şekil 2-1. Natriüretik peptidlerin eksprese olduğu dokular, proteoliz süreci,
matür protein hali. ... 3
Şekil 2-2. 17 amino asitten oluşan halka yapısı olan sistein-sistein disülfit çapraz bağı tüm natriüretik peptitlerde ortaktır . ... 4
Şekil 2-3. NPR-B reseptörünün homodimerik bölge yapısının şematik olarak gösterimi. CNP bağlanması sonrası intraselüler cGMP birikimi olmaktadır ... 7
Şekil 2-4. Natriüretik peptid reseptör topolojisi ve ligandları. ... 8
Şekil 2-5. Reseptörlerin aktivasyon ve inaktivasyon mekanizması ... 8
Şekil 2-6. Şematik olarak CNP/GC-B sistemi yolağının gösterimi ... 9
Şekil 2-7. FGF sinyali ve CNP/GC-B sistemi arası hücre içi etkileşiminin şematik gösterim. ... 11
Şekil 2-8. Kondrodisplazileri uzun kemiklerin (A-C) ve vertebranın (D, E) iskelet grafisi bulgularına göre sınıflama . ... 15
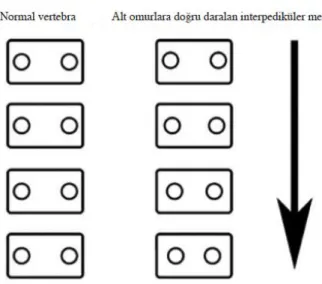
Şekil 2-9. İskelet displazilerinde saptanabilecek vertebra anormallikleri ... 55
Şekil 2-10. Vertebral pediküller arası mesafede azalmanın şematik görünümü... 56
Şekil 3-1. Standart solüsyon hazırlama görseli ... 77
Şekil 3-2. Sağlıklı Grup Polinom curve fit (<0,313 hariç) ... 80
Şekil 3-3. Hasta Grup Polinom curve fit (<0,313 hariç) ... 80
Şekil 4-1. Akondroplazili hastalarda ve sağlıklı grupta CNP konsantrasyon (ng/ml) değerlerinin karşılaştırılması ... 86
Şekil 4-2. Hasta ve sağlıklı grup CNP konsantrasyon (ng/ml) değerlerinin karşılaştırılması ... 86
Şekil 4-3. Akondroplazili hasta grupta cinsiyet ile CNP konsantrasyonu arası ilişki ... 87
Şekil 4-4. Hasta grupta cinsiyet ile CNP konsantrasyonu arası ilişki ... 87
Şekil 4-5. Kruskal Grup Kıyaslama Grafisi ... 89
Şekil 5-1. Akondroplazi’de FGFR3 gen mutasyonunun kemik gelişimi
üzerine etkisi ... 107
Şekil 5-2. FGFR3 sinyal yolağı ... 108 Şekil 5-3. Major FGFR3 mutasyonları topolojisi ... 111
KISALTMALAR
˚C :Santigrat
ACH : Akondroplazi
ACMG : American College of Medical Genetics and Genomics
ALS : Asit labil subunit
AMDM : Maroteaux tipi iskelet displazisi ANP : Atrial natriüretik peptid
AOI : Otozomal dominant perinatal ölümcül atelosteogenez I AOIII : Otozomal dominant öldürücü atelosteogenez III
Arg : Arjinin
ASD : Atriyal septal kusur
ATD : Asfiksik torasik displazi
AVP : Arjinin-vazopresin
VKİ : Vücut-kitle indeksi
BNP : Beyin natriüretik peptid
BOS : Beyin omurilik sıvısı
cGK : Siklik guanozin monofosfat-bağımlı protein kinaz cGKI : Siklik guanozin monofosfat-bağımlı protein kinaz I
cGKII : Siklik guanozin monofosfat-bağımlı protein kinaz II
cGMP : Siklik guanozin monofosfat
cm : Santimetre
CNP : C-tip natriüretik peptid
CNP-22 : C-tip natriüretik peptid-22 CNP-53 : C-tip natriüretik peptid-53
CNP-KO : CNP-nakavt
Col2-CNP-Tg : Spesifik CNP-transgenik fareler COMP : Kartilaj oligomerik matriks protein
dk : Dakika
DNP : Dendroaspis natriüretik peptit
ERK : Ekstraselüler sinyal ilişkili kinaz
ERT : Enzim replasman tedavisi
FGF : Fibroblast büyüme faktörü
FGF2 : Fibroblast büyüme faktörü-2
FGFR-3 : Fibroblast büyüme faktörü reseptörü-3
FLNA : Filamin A
FLNB : Filamin B
FLNC : Filamin C
GALNS : Galaktozamin-6-sülfat sülfataz
GC-A : Guanilat siklaz A
GC-B : Guanilat siklaz B
Gly : Glisin
GSK3β : Glikojen sentaz kinaz 3 beta
IGFBP : İnsulin benzeri büyüme faktörü bağlayıcı protein IGF-I : İnsulin benzeri büyüme faktörü-I
IGF-II : İnsulin benzeri büyüme faktörü-II
IL-1 : İnterlökin-1
IU : İnternasyonal ünite
IUGR : İntrauterin büyüme kısıtlılığı
Kg : Kilogram
LP : Little People Societies
LWD : Leri-Weill diskondrosteozisi
m2 : metrekare
MAP : Mitojen aktive edilmiş protein
MAPK : Mitojen aktive edilmiş protein kinaz yolağı
MED : Multiple Epifizyal Displazili
ml : Mililitre
mm : Milimetre
MPA : Mukopolisakaridoz
MW : Mann Whitney U testi
n : Sayı
ng : Nanogram
Nppc : CNP geni
NPR2 : Natriüretik peptid reseptör 2 NPR3 : Natriüretik peptid reseptör 3 NPR-A : Natriüretik peptid reseptör A NPR-B : Natriüretik peptid reseptör B NPR-C : Natriüretik peptid reseptör C
NTproBNP : N-terminal pro beyin natriüretik peptid NTproCNP : N-terminal pro C-tip natriüretik peptid
OD : Otozomal dominant
OI :Osteogenezis imperfekta
OR : Otozomal resesif
Ort : Ortalama
PAR1 : Psödootozomal bölge 1
proANP : Atrial natriüretik propeptid proBNP : Beyin natriüretik propeptid proCNP :Pro C-tip natriüretik peptid
PTH : Paratiroid hormonu
PTHR1 : Paratiroid hormon 1 reseptörü
PTHrP : G protein ilşkili PTH-PTH ilişkili peptid RMRP : Kıkırdak saç hipoplazisi
RMRP : Kıkırdak saç hipoplazisi
SADDAN : Gelişimsel gecikme ile giden ciddi akondroplazi ve akontozis
nigrikans
SCT : Spondilokarpotarsal sendrom
SDS : Standart deviasyon sapması
SED : Spondiloepifizyal displazi
SGA : Gestasyonel yaşa göre küçük
SHOX : Kısa boy homoebox içeren gen
STAT1 : Sinyal transdüser ve transkripsiyon aktivatörü
TD : Tanatrofik displazi
TNF-α : Tümör nekroz faktör alfa
TNF-β : Tümör nekroz faktör beta
TNSALP : Doku nonspesifik alkalen fosfataz
TSE : Türk Standartları Enstitüsü
USG : Ultrasonografi
VKİ : Vücut kitle indeksi
1 GİRİŞ VE AMAÇ
Büyüme, çocuğun sağlıklı olduğunu gösteren duyarlı bir göstergedir. Hücrelerin dinamik, heterojen, karmaşık bir süreç boyunca çoğalmasını ve farklılaşımasını içerir (1).
Çocukluk döneminde birçok hastalık, büyüme ve gelişmenin etkilenmesine yol açarak, boy kısalığına neden olmaktadır. İskelet displazileri büyük kısmı orantısız boy kısalığına yol açmaktadır (2).
İskelet displazisi; periostal kemikleşmenin normal olmasına karşın, enkondral kemikleşme bozulduğu, iskelette malformasyonların eşlik ettiği sendromların oluşturduğu heterojen bir gruptur. Ektodermal ve mezenkimal doku gelişimindeki kusur ya da dengesiz dağılım sonucu oluşmaktadır (3). İskelet displazilerinde boy kısalığı, hafiften ciddi dereceye kadar geniş yelpazededir (4).
İskelet displazileri; her ne kadar sık görülen hastalıklar olarak tanımlanmasa da insidansı 1/5000 civarında olup, iskelet displazilerine erken tanı koymak birçok morbiditenin önlenmesinde etkin olmaktadır (2).
200’ün üstünde farklı iskelet displazisi bulunmaktadır. Bu hastalarda gerek iskelet ile ilişkili gerekse iskelet dışında pek çok komplikasyon görülmektedir. Bunlardan %70’i perinatal dönemde letaldir. Kıkırdak-kemik dokuda eksprese olan mutant genler primer iskelet displazilerine yol açar. Metabolik kusur gibi kemik doku dışı faktörler iskelet gelişimini etkiler. Buna örnek olarak hipofosfatemik rikets gösterilebilir (3).
Büyüme genetik, beslenme, çevresel ve hormonal faktörlerden etkilenmektedir. Büyüme şekli ve erişkin olarak ulaşılacak boyun programlanmasında genetik faktörler en önemli rolü oynar. İkizlerde yapılan çalışmalar; %80 gibi büyük bir oranla, ulaşılacak boyun genetik faktörler tarafından programladığını göstermiştir. Boya etkili olduğu gösterilen birçok gen; büyüme kıkırdağında enkondoral kemikleşmeyi başlatmakta önemli rol oynamaktadır. Genom ilişkili yapılmış çalışmalar; insanlar arasında boy ile ilgili değişikliklerde rol oynayan genlerin natriüretik peptid sistemi üzerinde haritalandığını göstermiştir (1).
Natriüretik peptid ailesi; atrial natriüretik peptid (ANP) , beyin natriüretik peptid (BNP) , C-tip natriüretik peptid (CNP) ve dendroaspis natriüretik peptit (DNP)’den oluşmaktadır(1). Yapılan deneysel çalışmalar, CNP’nin kemik gelişiminde de rol oynadığını göstermektedir. Deneysel çalışmalarda CNP genindeki değişiklikliklerin "nakavt" farelerde ağır boy kısalığına yol açtığı gösterilmiştir (5). Akandroplazi, tanatrofik displazi ve hipokondroplazisi olan hastalarda plazma C-tip natriüretik peptid düzeyleri yüksek bulunmuştur (6). Hayvan modellerinde yapılan bir çalışmada tibia organ kültüründe CNP’nin kemikte uzamaya neden olduğu gösterilmiştir. CNP nakavt farelerin kısa boylu olduğu saptanmıştır. CNP’nin enkondral kemik oluşumunda etken olduğu ve CNP/GC-B sisteminin aktivasyonu sonucunda oluşan akandroplazinin tedavisinde kullanılabileceği ortaya konulmuştur (7).
İskelet displazisinin ayırıcı tanısı için detaylı anamnez, fizik muayene (özellikle antropometrik ölçümler), kemik survey, genetik incelemeler, ileri laboratuvar çalışmaları ve multidisipliner yaklaşım oldukça önemlidir. Bu çalışmanın amacı; kemik gelişiminde etkin olan C-tip natriüretik peptidin tüm iskelet displazi tipleri için spesifik bir belirteç olarak kullanılabilme olasılığını değerlendirmek, morbiditesi, hatta mortalitesi yüksek olan bu grup hastalığın tanısının daha erken konulabilmesini sağlamak ve tedavisi için ışık tutmaktır.
2 GENEL BİLGİLER
2.1 Natriüretik Peptidler
Atrial natriüretik peptid; ilk olarak Kisch tarafından 1956’da kalp atriumunda elektron yoğun cisimciklerin farkedilmesi ile bulunmuştur. 1979 yılında de Bold; bu bu granüllerin sayısının dehidratasyon sırasında azaldığını, tuz yüklemesinde ise arttığını göstermiş ve 1983 yılında sıçan atrial dokusundan bu natriüretik peptidi izole etmiştir. 1984 yılında ise bu peptid, Matsuo ve Kangawa tarafından insan atriumunda gösterilmiştir.
1988’de Sudoh ve arkadaşları domuz beyninden B-tip natriüretik peptit (BNP) ve 1990 yılında C-tip natriüretik peptit (CNP) izole etmişlerdir. ANP ve BNP sentezinin yapıldığı organla birlikte anılmış; kardiyak hormonlar olarak isimlendirilmişlerdir. 1992 yılında Shweitz ve arkadaşları yeşil mamba yılanının zehrinden dendroapsis natriüretik peptiti (DNP) izole etmiştir (5).
ANP, BNP, CNP dokularda pro-hormonlar olarak eksprese edilir (Şekil 2-1). ANP, korin tarafından oluşturulurken; BNP ve CNP'nin oluşumundaki görevli enzim henüz bilinmemektedir. Tüm natriüretik peptidlerin disülfit bağları ortaktır (Şekil 2-2).
Şekil 2-2 17 amino asitten oluşan halka yapısı olan sistein-sistein disülfit çapraz bağı tüm natriüretik peptitlerde ortaktır (5).
2.1.1 ANP
Kardionatrin, atriopeptin ve atrial natriüretik peptid (ANP) gibi birçok farklı isimle anılan ANP; başlıca kalpten salınır (8). Bunun dışında hipotalamus, hipofiz, adrenal medulla, gastrointestinal sistem, timus, korpus luteum, ovaryum, testis gibi organlardan da salınır (5).
Preprohormon olarak sentezlenen ANP; 151 aminoasit uzunluğunda olup, aminoterminal bölümü ayrılması sonucu 126 aminoasitli proANP oluşur. ProANP şeklinde depolanır (8).Sisteme salınacağında ise atriopeptidaz ile proANP 98 ve 28 aminoasitten oluşan iki parçaya ayrılır (5).
ANP'nin plazma düzeyi sağlıklı bireylerde yaklaşık olarak 10 fmol/ml olarak tanımlanmıştır. Konjestif kalp yetmezliği olan bireylerde ise 10-30 kat artmış olarak bulunur. İnsan ANP geni; kromozom 1p36.2 üzerinde lokalizedir. ANP geni ; BNP, CNP gibi 3 ekzon bölgesi içerir (8). Plazmada moleküler ağırlığı 3085,5 Dalton olmakla birlikte, normal insandaki ANP konsantrasyonu ortalama 0,65 ng/l civarıdır (5).
ANP primer olarak atrial granüllerde depolanır ve eksprese edilmekle birlikte; düşük konsantrasyon da olsa ventrikul ve böbrek gibi dokularda da bulunur. ANP salınımı için birincil uyarıcı intravaskuler volum artması sonucu atrium duvarının gerilmesidir. Sekrete edilen ANP ilk olarak koroner sinuse perfüze olur; daha sonra buradan hedef organlara dağılır. Endotelin, anjiyotensin, arginine-vazopresin (AVP) ; ANP salınımını uyararak böbreklerde natriüretik etki göstermesine neden olurlar (8).
2.1.2 BNP
B-tip natriüretik peptid (BNP) ilk olarak domuz beyninde tanımlanmıştır. Her ne kadar beyin natriüretik peptid olarak anılsa da en fazla kalpte bulunur. 134 aminoasitli prehormon olarak sentezlenip; 108 aminoasitli prohormon haline dönüşür. Henüz tanımlanmamış olan bir proteaz ile aktif protein haline dönüşür. 32 amino asitlik yapı pro-BNP’nin C ucuna, 76 amino asitli NT-pro-BNP ise N ucuna denk gelir. Aktif hali kanda dolaşan formudur (5).
İnsan BNP geni; ANP geninden 8kb yukarı yönde, kromozom 1p36.2 üzerinde lokalizedir (8). ANP ile birlikte atrial granüllerde depolanan BNP; kardiyak duvar gerilmesi sonucu ventriküllerden salınır (8). Hem ANP hem de BNP kardiyovasküler sistem üzerinde hemodinamik, diüretik, natriüretik, vazodilatator, anti-proliferatif ve anti-hipertrofik etkilere sahiptir (1).
Normalde plazma konsantrasyonu yaklaşık olarak 3.5 pg/ml ya da ANP’nin onda biri kadardır. Konjestif kalp yetersizliği, miyokard infarktüsü durumunda plazma BNP konsantrasyonu normal seviyesinden 200-300 kat yükselir. BNP; bu yüzden önemli bir kardiak gösterge olarak kullanılmaktadır (8).
2.1.3 CNP
CNP, natriüretik peptid ailesinin üçüncü üyesi olup, önemli natriüretik özelliği olmamakla birlikte endokondral kemik gelişiminin düzenlenmesinde anahtar rol oynamaktadır. CNP, birçok dokuda (kıkırdak, kemik, beyin, endotel, düz kas ve kalp sentezlenebilmesine rağmen; en çok büyüme plağının hipertrofik zonununda sentezlenmektedir (1,8).
Lineer büyüme için güçlü bir pozitif düzenleyicidir. CNP düzeyleri plazmada ölçülebilir. CNP; hücrelerden kendisi ile eşit mol oranıyla salınan amino terminal propeptit (NTproCNP)’ten sentezlenmektedir (6).
İlk olarak domuz beyninden elde edilmiştir. İnsan beyninde de CNP varlığı gösterilmiş olup, vasküler endotelyal hücreler, makrofajlar tarafından da salgılanmaktadır. Nöropeptid ve otokrin/parakrin regülator olarak rol oynadığı düşünülmektedir (9).
103 aminoasitli insan proCNP’nin yıkılması ile CNP-53; oluşup şu an için tanımlanamamış bir enzim tarafından yıkılarak CNP-22 meydana gelir. Her ikisinin
fonksiyonu benzer olsa da bulundukları yerler farklılık gösterir. CNP-22 plazma ve beyin omurilik sıvısında daha fazla bulunurken CNP-53 beyin, endotel hücresi, kalpte daha fazla miktarda bulunmaktadır (5).
CNP geni 2q24 ile 2q terminal kısmı arasında olup; Kromozom 1 üzerinde lokalizedir (8).
ANP ve BNP’den farklı olarak sentezi sonrası hemen sekrete edilir, depolanmaz. Endotel hücrelerinden TNF-α ,TNF-β, IL-1 ve süregelen stres vasıtasıyla salınır. Sentezi insulin tarafından inhibe edilir. Endotelden salınan CNP; kartilaj matriks sentezini başlatır, kondrosit proliferasyonunu ve farklılaşmasını stimüle eder (1,8).
2.1.4 DNP
DNP; natriüretik ailesinin en yeni tanımlanan natriüretik peptiddir. 38 amino asitten oluşur. ANP ve BNP gibi natriüretik peptit reseptör A (NPR-A) aktivitesine sahiptir (5).
DNP’nin vazodilatasyona yol açtığının bilinse de fonksiyonu konusunda oldukça az bilgi mevcuttur (1).
Konjestif kalp yetmezliği gibi durumlarda plazmaki DNP seviyesi yükselir. Deney hayvanlarıyla yapılan çalışmalarda natriüretik ve diüretik özellikleri gösterilmiştir (5).
2.1.5 Osteokrin/ Musklin
Natriüretik peptid ile benzerlik gösteren iki grup peptid daha tanımlanmıştır. Birinci grup; ilk olarak kemikte bulunduğu için osteokrin olarak adlanırılırken, ikinci grup ilk olarak kasta bulunduğundan musklin olarak adlandırılmıştır. 2004 yılında “American Society of Bone and Mineral Research Conference”ında osteokrinlerin ANP ile kompetitif olarak NPR-C'ye yüksek affinite ile bağlandıkları açıklanmıştır. NPR-A ve NPR-C aynı hücrede eksprese edildiği zaman osteokrin; NPR-C ilişkili ANP degradasyonunu inhibe ederek ANP-bağımlı cGMP artışına yol açar. Fare modellerinde yapılan osteokrinin transgenik eksprese olduğu çalışmalar neticesinde, osteokrinin CNP'nin NPR-C'ye bağlanmasını bloke ederek büyüme plağındaki CNP seviyelerini artırdığını düşündürmektedir (8).
2.2 Natriüretik Peptid Reseptörleri
Natriuretik peptitlerin 3 tip reseptörü bulunmaktadır (5): Natriüretik Peptid Reseptör A (NPR-A)
Natriüretik Peptid Reseptör B (NPR-B) Natriüretik Peptid Reseptör C (NPR-C)
Natriüretik peptidlerin biyolojik etkisi üç reseptör subtipine bağlıdır. Bunlardan ikisi homodimerik reseptör olan NPR-A/GC-A/NPR1 ve NPR-B/GC-B/NPR2’dir. Bu iki reseptör transmembranöz guanilat siklaz vasıtası ile cGMP sentezini katalizler. Üçüncü reseptör ise NPR-C/NPR3’dir. Bu reseptörün intirinsik enzimatik aktivitesi yoktur. Natriüretik peptidlerin lokal konsantrasyonunu yapısal reseptör aracılı internalizasyon ve degradasyona yol açarak kontrol eder (1,8).
ANP ve BNP, GC-A aracılığı ile kardiyavasküler sistemde homeostaz regülasyonunda temel rol oynar. Kalp yetmezliği, miyokard infarktüsü, kardiak hipertrofi ve hipertansiyon gibi çeşitli patofizyolojik stres sırasında kandaki seviyeleri artar. ANP ve BNP; primer olarak atriyum ve ventrikülden salınan güçlü diüretik, natriüretik, vazodilatatör etkileri olan peptidlerdir. ANP ve BNP kalp yetmezliği tedavisinde; kardiak yetmezlik ve kardiak hipertrofi belirteci olarak kullanılır (9).
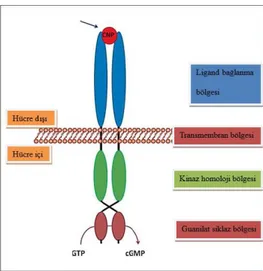
Şekil 2-3 NPR-B reseptörünün homodimerik bölge yapısının şematik olarak gösterimi. CNP bağlanması sonrası intraselüler cGMP birikimi olmaktadır (1).
CNP'nin reseptörü; natriüretik peptid reseptörü-B (NPR-B, genNPR2) olarak anılır. NPR-B; ikincil haberci olarak cGMP kullanan bir membran reseptörüdür (6).
NRP-B; ANP, BNP, CNP bağlandığı ekstraselüler ligand bağlanma bölgesi, transmembran bölgesi, intraselüler kinaz homoloji bölgesi ve natriüretik peptid stimulasyonu sonucu GTP’den cGMP’nin üretildiği karboksi-terminel guanilat siklaz bölgesinden oluşur (Şekil 2-3, Şekil 2-4). Reseptörler fosforilizasyon ile aktive olurken, defosforilazasyon ile inaktive olur (Şekil 2-5). Natriüretik peptidler enkondral kemik kemikleşmesinde (CNP, BNP) ve kardiyak sistem fonksiyonu üzerinde (BNP, ANP, CNP) NPR-B’ye bağlanarak farklı etkilerini gösteririler (10).
Şekil 2-4 Natriüretik ppetid reseptör topolojisi ve ligandları (8).
Şekil 2-5 Reseptörlerin aktivasyon ve inaktivasyon mekanizması (8).
Farelerde CNP/B sistemi ilişkili yapılan genetik araştırmalar; CNP ve GC-B’nin endokondral kemik gelişiminde esas rol oynadığını göstermiştir (9). cGMP bağımlı protein kinaz (cGK); natriüretik peptid ailesi ve guanilat siklaz sisteminin aktivasyonu sonucu oluşan son ürün olarak tanımlanmıştır. cGK geninin bir alt tipi
olan bu genin hasarlandığı fareler, cGKII (cGKII-KO fareler), kısa boy fenotipindedir. cGKII; CNP/GC-B sistem yollağı vasıtası ile enkondral kemik gelişimini etkiler. cGKII, GSK3β’yı fosforile ederek inaktive ederek β-katenin fonksiyonunu ve miktarını artırarak büyüme kıkırdağının hipertrofik diferensiyasyonuna yol açar (Şekil 2-6).
Şekil 2-6 Şematik olarak CNP/GC-B sistemi yolağının gösterimi (9).
Tip II kollajen kontrolu altındaki kartilaj spesifik CNP-transgenik fareler (col2-CNP-Tg fareler)’de endokondral kemikleşme ile kemiklerin aşırı büyüdüğü gözlenmiştir. Öteyandan CNP/GC-B-KO farelerde col2-CNP-Tg farelerin büyüme plaklarındaki her bir kondrosit tabakasının daha geniş olduğu gösterilmiştir. CNP/GC-B sisteminin enkondral kemik gelişiminde etkisi; CNP geni (Nppc) nakavt farelerde (CNP-KO fareler) bozulmuş kemik gelişimi nedeniyle kısa boy gözlenmiştir. Kemikler endokondral kemikleşme ve membranöz kemikleşme olmak üzere farklı iki mekanizma ile şekillenir. Kemiklerin çoğu; büyüme plağındaki proliferasyon, hipertrofi, hücre ölümü ve osteoblastik replasyon ile giden endokondral kemikleşme ile şekillenir. Kısa boy fenotipi gözlenen CNP-KO farelerde bu durumun endokondral kemikleşmedeki kusur ile kemik gelişiminin bozulması sonucu olduğu gösterilmiştir. CNP-KO farelerin büyüme kıkırdağının histolojik analizi, büyüme plağının her bir kondrosit tabakasının CNP-KO farelerde daha dar olduğunu kanıtlamıştır. Bununla birlikte GC-B geni (NPR2) hasarlanmış CNP-KO farelerde aynı kısa boy fenotipi gözlenmiştir. Bu da CNP/GC-B sisteminin fizyolojik olarak endokondral kemik gelişimindeki önemini kanıtlar niteliktedir. cGKII nakavt farelerde; bozulan enkondral osifikasyonundan ötürü kısa boy
gelişmesi; cGKII’nin CNP ilişkili enkondral kemikleşmede etkin olduğunu göstermiştir (9).
C-tip natriüretik peptid (CNP), iskelet gelişiminde önemli bir düzenleyicidir. CNP reseptör natriüretik peptid reseptör-B (NPR-B, gen NPR2) fonksiyon kaybına yol açan mutasyonlar boy kısalığına neden olmaktadır. Otozomal resesif tipte geçen tipleri genellikle akromezomelik displazi, Maroteaux tipte iskelet displazilerine yol açmaktadır. NPR2 geni heterozigot mutasyonları kısa boy ile ilişkilendirilmiştir (11). Ek olarak, homozigot ya da bileşik heterozigot NPR2 mutasyonları; oldukça kısa boy ile karşılaşılan olan Maroteaux tipi iskelet displazisine (AMDM; OMIM * 602875) yol açmaktadır. NPR2 geninde fonksiyon kaybı ile sonuçlanan heterozigot mutasyonların idiyopatik kısa boya neden oldukları gösterilmiştir (6). Her 700 insandan birinin bilinmeyen şekilde NPR2 mutasyonunu taşıdığı göz önüne alındığında 30 kişinin birinde görülen idiyopatik kısa boy fenotipinin nedeninin NPR2 mutasyonu taşıyıcılığı olabileceği düşünülmektedir (9). Buna karşılık, fonksiyon kazanımı ile sonuçlanan NRP2 genindeki heterozigot mutasyonların uzun boya neden olduğu gösterilmiştir. Dengeli translokasyonla, CNP overekspresyonu sonucunda benzer fenotipik özellikler bazı bireylerde gözlenmiştir (1). Yapılan çalışmalarda; AMDM'li bir çocukta CNP ve NTproCNP'nin plazma konsantrasyonlarının yüksek olduğu bildirilmiştir. Bu da azaltılmış hücre içi CNP aktivitesinin sonucu CNP üretimini artırabileceğini düşündürmektedir (6).
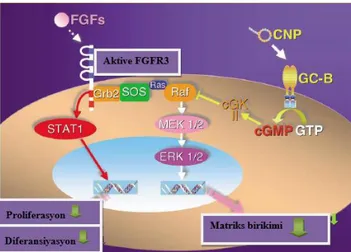
Akondroplazi; iskelet displazilarinden en sık görüleni olup, kısa uzuvlu boy kısalığı karakterize olup, FGF reseptörü 3 (FGFR3) geni yapısal mutasyonu sonucu oluşmaktadır. Yapılan çalışmalar; CNP’nin akondroplazili hastalarda tedavi edici nitelikte olduğunu göstermektedir. FGFR-3, büyüme plağındaki birçok kaskadı aktive eder. CNP’nin akondroplaziyi iyileştirdiğini açıklığa kavuşturmak için CNP’nin FGFR3 biyolojik etkisine aracılık eden ERK sinyali üzerine etkisini incelemek gerekir. Bu kaskadlardan en önemli yollağı; kondrosit proliferasyonunu inhibe eden STAT1 yolağı ve kondrosit proliferasyonunu önleyen, yapı bozulmasını artıran MAPK kinaz (MEK)/ERK MAPK yolağıdır. Bu iki yolak kondrositik farklılaşmayı engelleyip, matriks bozunmasını arttırırak kemik gelişiminin yetersizliğine neden olur. MEK/ERK MAPK yolağı ve CNP hücre içi yolağı birbirlerinin inhibitörleridir. CNP, FGF2 stimulasyonu ile fosforile olan ERK’i; GC-B ligasyonu sonucu cGMP aktivasyonunu sağlayarak doz bağımlı olarak inhibe eder. Bunun sonucunda kondrositler tarafından matriks sentezi artar. Kondrosarkomlu fare
modeli kondrositleri kullanılarak yapılan invitro çalışma; CNP’nin, FGF sinyali ERK yolağını; cGKII aktivasyonu ile Raf-1 seviyesinde inhibe ettiğini göstermiştir (Şekil 2-7). FGFR-3’ün aşırı çalışması sonucu NRP-B’nin fonksiyonunun inhibe olur. NPR2 geninde fonksiyon kaybına yol açan mutasyon ile CNP plazma seviyesinin artmış olması; FGFR-3’ün yapısal aktivasyonu sonrası oluşan hastalıklarda da CNP düzeyinin artacağını düşündürmüştür (8,9).
Şekil 2-7 FGF sinyali ve CNP/GC-B sistemi arası hücre içi etkileşiminin şematik gösterimi (9) Üçüncü natriüretik peptid reseptörü, temizleme reseptörü olarak adlandırılan C-reseptörü (clearance receptor)’dür. Reseptör ilişkili natriüretik peptid degradasyonunda rol oynamaktadır (9).
NPR-C; intraselüler guanil siklaz bölgesi olmayan natriüretik peptid reseptörüdür. Natriüretik peptidlerin reseptör aracılı yıkımından sorumludur. Natriüretik peptidler ayrıca endopeptidazlar aracılığıyla da yıkılır (1). NPR-A ve NPR-B membran bağımlı reseptörler olup; ekstraselüler ligand bağlanma bölgesi, tek hidrofobik transmembran bölgesi, intraselüler kinaz homoloji bölgesi, dimerizasyon bölgesi, karboksi-terminal guanilat siklaz bölgesi içerir.
C'nin ekstraselüler ligand bağlayıcı bölgesi ; yaklaşık %30 oranda NPR-A ve NPR-B'ye benzer. NPR-NPR-A ve NPR-B'den farklı olarak 37 intraselüler aminoasit içerir. NPR-C şeklindeki yatay kırmızı çizgi (Şekil 2-4) intermoleküler disülfat bağını göstermektedir (8).
Önceden belirtildiği gibi C-reseptörlerinin natriüretik peptid yıkımına neden oldukları düşünülmektedir. C-reseptörü hasarlanmış farelerde aşırı büyüme ile sonuçlanan iskelet gelişiminin col2-CNP-Tg fareler gibi olduğu gözlenmiştir.
Osteokrinin fazla eksprese olduğu transgenik farelerde ise iskeletin aşırı gelişiminin osteokrinin C-reseptör için endojenik ligand olmasından kaynaklı olduğu düşünülmektedir. CNP’nin azalmış yıkımı, CNP’nin büyüme plağındaki miktarının artmasına yol açarak; artmış CNP’nin endokondoral kemik gelişiminin uyarılmasına yol açtığını düşündürmüştür (9). (Şekil 2-6)
2.3 Büyüme Üzerine Etkili Diğer Hormonlar
Tüm dokularda protein sentezini artıran büyüme hormonu en belirgin etkisini iskelet sistemi üzerinde gösterir. Bunu kondrosit ve osteojenik hücrelerde protein depolanmasını artırarak, kondrosit ve osteojenik hücrelerin proliferasyonunu artırarak ve kondrositlerin osteojenik hücrelere dönüşümünü sağlayarak yaparlar. Bu etkilerini insulin benzeri büyüme faktörleri (somatomedinler) tarafından gerçekleştirirler. İnsulin benzeri büyüme faktörleri; büyüme hormonları aracılığı ile başlıca karaciğerde sentezlenir. Dört somatomedin arasında en değerlisi somatomedin C olarak da bilinen insulin benzeri büyüme faktörü-I (IGF-1)’dir (12). Büyüme hormon etkilerini başlıca IGF-I ve onun bağlayıcı protein IGFBP-3 aracılığıyla gösterir. Ergenlik döneminde seks steroidlerinin artışına paralel olarak IGF-I de artmaktadır. Tiroid hormonları ya doğrudan kemik üzerine etkisini göstererek büyümeyi sağlar ya da büyüme hormon- IGF-I aksı üzerinde etkisini gösterir. Glukokortikoidler ise hem büyüme hormonunu baskılayarak hem de IGF-I’in işlevini azaltarak büyümeyi negatif etkiler (13).
Postnatal dönemde büyüme tiroid hormon ve büyüme hormon vasıtası ile sağlanırken, fetal dönemde insulin benzeri büyüme faktörleri (IGF-I, IGF-II) aracılığıyla büyüme sağlanır. Fetal dönemde IGF-II, IGF-1’e göre daha potenttir (13).
IGF’ler insulin benzeri büyüme faktörü bağlayıcı proteinlere (IGFBP’ler) bağlanarak taşınır. IGF-I’in temel hedef organı büyüme kıkırdağı olup, hipofize negatif geri besleme ile büyüme hormon sentezini azaltır. Büyüme hormon fazlalığında IGF’ler ve bağlayıcı proteinleri artarken, eksikliğinde azalırlar. IGF’ler yapı olarak insuline benzer. IGF-I 12. Kromozom uzun bacağında yer alırken, IGF-II 1. kromozomun kısa bacağında yer alır. IGF-I ve IGF-II hem invivo hem de invitro olarak büyüme üzerine pozitif etkilidir. Reseptörler üzerinden etkilerini gösterirler. Tip 1 ve tip 2 olmak üzere iki tip reseptörleri vardır. Tip 1 reseptörün IGF-I’e karşı
afinitesi fazladır. İki subunitten oluşur. Bunlar α ve β subunitiir. Reseptörün intraselüler tirozin kinaz aktivitesi bulunur. İnsulin benzeri büyüme faktörü bağlayıcı proteinler altı sınıfa ayrılır. IGFBP-1; amniyon sıvısında büyük oranda bulunur. IGFBP-2 beyin omurilik sıvısı (BOS)’nda, seminal plazmada bulunur. IGFBP-3 en önemli bağlayıcı protein olup serumda bulunmaktadır. IGF’ler asit labil subunit (ALS) ile birlikte IGFBP’lere üçlü karmaşık olarak bağlanıp serumda bulunurlar. IGFBP-4 serum ve seminal sıvıda, IGFBP-5 BOS ve az miktirda serumda bulunur. IGFBP-6; transforme fibroblastlar tarafından üretilmektedir, BOS’ta bulunur. IGFBP’ler trofik hormonların etkisi ile IGF ve resptörleri arası ilişkiyi düzenler (14).
2.4 İskelet Displazileri (Osteokondrodisplaziler)
2.4.1 Giriş
Kas iskelet sistemi; kemikler, eklemler, ligamanlar, kaslar ve bağ dokusundan oluşmaktadır. Doğumsal kemik hastalıkları çoğunluğu kalıtsal olmakla birlikte; iyatrojenik olarak aminopterin,talidomid, fenitoin, trimetadon gibi teratojen ajanlar doğumsal iskelet deformasyonlarına neden olabilir.
Süt çocukluğu döneminde ve sonraki yaşlarda görülen iskelet hastalıkları en sık travma ve buna bağlı fraktürler nedeniyle oluşur.
Kemiklerin dejeneratif ve vasküler hastalıkları ile çok sık karşılaşılmamakla birlikte bu grupta epifizin vaskülarizasyon bozukluğu sonucu ortaya çıkan osteokondrozlar yer alır. Hemangiom ve lenfanjiektazinin bulunduğu ekstremiteler diğerlerine göre daha büyük olabilir.
Kronik kurşun zehirlenmesi, metabolik bozukluklar, beslenme bozuklukları da kemik değişikliklerine neden olabilir. Örneğin; A vitamininin uzun süre aşırı dozlarda verilmesiyle özellikle uzun kemiklerin diafizlerinde belirgin olarak kortikal hiperostoz oluşmakta; C vitamini eksikliğinde ise metafizde kalın beyaz çizgi, epifizde buzlu cam görünümü, korteks incelmesi, subperiostal hematomlar izlenir. D vitamini eksikliğinde uzun kemiklerin uç kısımlarının düzensizlik gözlenir. Kortekste demineralizasyon, kemiklerde eğrilmeler, kostokondrial birleşme yerlerinde tesbih dizisi şeklinde şişlikler izlenebilir (15).
2.4.2 Normal Kemik Gelişimi
Kemik; mezenkimal hücrelerin yoğunlaşması ile oluşmaktadır. Hiperplastik olan kartilajinöz doku orta kısımda yer alır ve hücre dışı matriks mineralize olur. Kan damarlarının oluşumu ve osteoblastların bu bölgeye göçü ile endokondral kemikleşme gerçekleşir. Bu bölge birincil kemikleşme merkezidir. Epifiz bölgesinde benzer evrelerle kemik dokusu oluşur; bu bölge sekonder kemikleşme merkezi olarak anılır. Bu iki kemikleşme merkezi arasındaki kıkırdak dokusu bulunur ve büyüme plakası (fizis) olarak adlandırılır. Düz kemikler, mezenşimal hücrelerin doğrudan osteoblastlara dönüşmesi (intramembranöz kemikleşme) ile meydana gelir (16).
2.4.3 İskelet Displazileri Sınıflandırılması
İskelet displazisi (osteokondroplazi) kemik, kıkırdak veya her iki dokuyu etkileyen, kısa boy ile karakterize geniş heterojen bir genetik hastalık grubudur (16). Ektodermal ve mezenkimal doku gelişimindeki kusur ya da dengesiz dağılım sonucu oluşmaktadır. Şimdiye kadar tanımlanmış 200’den fazla iskelet displazisi mevcuttur. Bunlardan %70’i perinatal dönemde letaldir. Organogenez sırasındaki sinyal kusuru sonucu bu hastalık grubu oluşmaktadır. Spesifik kusurlar bazı hastalıklarda görülmektedir. Örneğin; Holt Oram Sendromu’nda izole polidaktili, birinci metakarpta kısalık görülmektedir. Sinyal kusuru tek başına bir bölgeyi etkileyebildiği gibi pek çok organı bir arada etkileyebilmektedir. Prenatal süreçte eksprese olan genler, postnatal süreçte de eksprese olmaya devam ederse displazi oluşur. Kıkırdak-kemik dokuda eksprese olan mutant genler primer olarak iskelet displazilerine yol açar. Metabolik kusur gibi kemik doku dışı faktörler iskelet gelişimini etkiler. Buna örnek olarak hipofosfatemik rikets gösterilebilir (3).
Değişik özellikleri olan bu hastalıkları gruplamak için uluslarası sınıflama ve terimlendirme ise ilk olarak 1969 yılında yapılmış olup, daha sonra güncellenerek 1977, 1983, 1991, 1997,2001, 2005, 2010, 2015 yılında yapılmıştır (4).
Terminolojide ‘’boy kısalığı’’ deyimi yerini kemikte büyüme ve gelişme bozukluğunu yansıtan ‘’displazi’’ terimine bırakmıştır. İskelet displazileri, boy kısalığı nedenleri arasında orantısız boy kısalığı nedenleri arasındadır (15). Ailesel boy kısalığı ve yapısal boy kısalığı ekstremitelerde ve gövdede simetrik kısalık ile
görülür. Öteyandan infant ve çocukların takibinde dismorfik ve/veya çok sayıda konjenital anomali eklenebilir (3).
Bazı iskelet displazileri kısa ekstremite ile kendini gösterirken [akondroplaziler, kondroektodermal displaziler, metafizer kondrodisplaziler, trikorinofalangeal sendrom gibi) bazıları belirgin olarak kısa gövde (metatropik displazi, Kniest sendromu, spondiloepifizer displazi sendromları, mukopolisakkaridoz sendromları (I,II,IV,VI)] ile karakterizedir (15).
İskelet displazileri; kemik veya kıkırdak dokunun büyüme ve gelişmesinin bozulduğu hastalık grubudur. Aşağıda sıralanan üç grupta incelenebilir;
1)Kemiğin büyüme bozuklukları (vertebra, pelvis ve uzun kemiklerin metafiz ya da epifizleri etkilenmiştir.)
2)Kıkırdak doku veya fibröz doku organizasyonunda düzensizlik 3)Kemiğin şekil veya yoğunluk değişiklikleri
Kemik lezyonlarının radyolojik lokalizasyonuna göre kabaca aşağıdaki gibi sınıflanır (Şekil 2-8) (4);
1. Normal (A+D) 2. Epifizyel (B+D) 3. Metafizyel (C+D) 4. Spondiloepifizyal (B+E)
5. Spondiloepimetafizyal displazi (B+C+E)
Şekil 2-8 Kondrodisplazileri uzun kemiklerin (A-C) ve vertebranın (D, E) iskelet grafisi bulgularına göre sınıflama (4).
Bu hastalık grubu; klinik, laboratuvar, radyolojik ve genetik açıdan pek çok alt sınıfa ayrılmaktadır. Toplum arasında hastalık grubunun insidansı ortalama 1/3000-5000 arasındadır (16). 1970’ten önce iskelet displazisi düşünülen hastalar iki sınıfa ayrılmıştır. Hastalar kısa uzuvlu ise akondroplazi, kısa gövdeli hastaların ise
Morquio Sendromu tanısı almışlardır (4). İskelet displazilerinin değerlendirilmesinde başlangıca göre kıyaslandığında uzun ve iyi bir yol katedilmiştir. 1970 yılında, iskelet displazilerine ait ilk uluslararası sınıflandırılma yayınlanmıştır (17).
Bu alandaki biyokimyasal ve genetik gelişmelere paralel 100 farklı iskelet hastalığı klinik ve radyolojik özelliklerine göre sınıflandırılmış ve hastalıkların anlaşılması kolaylaşmıştır. Moleküler genetikteki son gelişmeler ile çok sayıdaki iskelet displazilerine neden olan temel kusurlar tanımlanabilmiştir. 5. Uluslararası Terminoloji Komitesi (The fifth International Nomenclature Committe) 1997’de Los Angeles’ta sadece terminoloji güncellemekle kalmamış yeni tanımlanmış çok sayıdaki sendrom da sınıflamaya eklenmiştir. 2001 ve 2005 yıllarında klinik ve patojenik mekanizma tekrar gözden geçirilmiş; sınıflamada kullanılmıştır. Böylece klinik, radyolojik, morfolojik ya da biyokimyasal özelliklere göre ortak patogenezi olan hastalıklar bir grupta toplanmıştır. Bu hastalıkların çoğu hem iskelet ile ilgili hem de iskelet dışında birçok komplikasyona yol açmaktadır. İskelet displazilerine verilen isim ya etkilenen bölgeye (epifizyel displazi, metafizyel displazi gibi) göre verilmiştir ya da Yunan terminolojisine göre konulmuştur. Bazı hastalıklar ise adını veren kişiler (Kniest displazi, Ellis-Von Cleveland Sendromu gibi) ile anılmaktadır. Bazen de patogenezi tanımlama için (osteogenezis imperfekta gibi) isimler derive olmuştur ama genellikle yanlış kullanımla (akondroplazi ve akandrogenezis gibi) sonuçlanmıştır (4).
Uluslarası sınıflamaya göre eskiden kabaca iskelet displazileri 5 major gruba ayrılırdı (15).
1. Osteokondroplaziler: Kartilajdaki ya da kemik büyüme-gelişmesindeki anomali.
2. Dizostozlar: Tek başına ya da birkaç kemiğin kombinasyonu ile oluşan malformasyon (iskeletin jeneralize hastalığını yansıtmaz).
3. İdiyopatik osteolizis: Kemiğin multifokal rezorpsiyonu ile giden bir grup hastalık.
4. Kromozomal aberasyon ile ilşkili iskelet hastalıkları 5. Primer metabolik hastalık ile ilişkili iskelet hastalıkları
Osteokondroplaziler boy kısalığı ayırıcı tanısında önemli bir grubu oluşturur. Bu karmaşık grup hastalıklar kartilajın primer anomalileri ile veya kemik büyüme-gelişmesindeki anomaliler ile ilişkilidir. Bu grup aşağıdaki gibi sınıflandırılmıştır (15).
1. Tübüler kemik ya da vertebral kemik büyümesinde veya her ikisinde olan kusur. ( Kondrodisplazi olarak da adlandırılabilir.)
Kemiğin dansitesi, miktarı ve remodulasyonunda anomali (kemikteki yoğunluk artışı ya da azalışı, mineralizasyon ve mineral metabolizmasındaki bozukluğu içerir.)
Kartilaj ve fibroz bağ dokusunun gelişimi esnasında disorganizasyonunu içeren hastalıklar 2001 ve 2005 yıllarında klinik ve patojenik mekanizma tekrar gözden geçirilmiş; klasifikasyonda kullanılmıştır. Böylece klinik, radyolojik, morfolojik ya da biyokimyasal özelliklere göre ortak patogenezi olan hastalıklar bir grupta toplanmıştır (15).
Yıllar boyunca çok sayıda revizyon yapılmıştır ve son sınıflandırma 2015 yılında yayınlanmıştır. Genetik İskelet Bozukluğunun Nozolojisi ve Sınıflaması (18); gen ailelerine (FGFR3 grubu), fenotipik sunuma (akromikrik displazi) ve patofizyolojiye (lizozomal bozukluklara) dayalı 42 gruba ayırır. Bu sınıflamada 364 genin tarif edildiği 436 bozukluk bulunur. Bazı tipler için henüz bir genetik etiyolojinin bulunmadığını belirtilmiştir (Ek.1). Bununla birlikte, bir gen birden fazla hastalık ile ilişkili olabilir. Örneğin; COL2A1 mutasyonları akondrojenez tip 2, hipokondrojenez, spondiloepifizyel displazi, Kniest ve Stickler sendromlarına yol açabilir. Bir hastalığa birden çok mutasyon yol açabilir. Örneğin; SLC26A2, COMP, MATN3, COL9A1, COL9A2 ve COL9A3'teki mutasyonlar epifizyel displaziye yol açmaktadır (17).
İskelet displazisi olan hasta grubunda çeşitli konjenital malformasyonlar, spinal anomaliler, hipoglisemi, hipo/hiperkalsemi, adrenal yetmezlik, pubertal gelişim sorunları ve cinsiyet farklılaşması bozuklukları gibi endokrinolojik problemler de görülebilir (16).
Bu gruplar semptomların başlama yaşı ve ilişkili klinik anomalilere göre alt gruplara ayrılmıştır. Bazı hastalıklar ise kalıtım şekillerine göre (dominant ya da X’e bağlı geçen spondiloepifizyel displaziler gibi) gruplanmıştır (4).
En yaygın olarak kullanılan sınıflama ise radyolojik olarak saptanan anomalilere göre olan sınıflamadır. Uzun kemiklerdeki anomalinin bulunduğu segmente göre (epifiz, metafiz, diafiz) sınıflama yapılır. Vertebranın etkilenmesine göre ise spondiloepifizyal ya da spondilometafizyel displazi olarak adlandırılır. Yine bu sınıflamalar hastalıkların klinik ve radyolojik özelliklerine göre alt gruplara ayrılır. Bu hastalıkların genellikle patogenezi, morfolojisi, biyokimyasal ve moleküler kusuru çözümlenmiştir. Terimlendirme spesifik patogenez veya metabolik
kusura göre değişmektedir. Etyolojik veya patolojik terimlendirme bazı iskelet displazileri [mukopolisakkoridozlar, mukolipidozlar (β-glukuronidaz eksikliği, fukosidoz gibi), mineralizasyon bozuklukları (hipofosfotazya gibi)] tarafından kullanılmaktadır (4).
2.4.4 Kemikte Büyüme Bozukluğu İle Karakterize Olan İskelet Displazileri
2.4.4.1 Tanatrofik Displazi
Tanatrofik displazi (TD) genellikle yenidoğan döneminde; küçük göğüs kafesi, akciğer hipoplazisi, solunum yetersizliği nedeniyle solunum baskılanması sonucu letaldir (19).
Hastalık çoğunlukla perinatal dönemde tanı alır. Hastalık letaldir. Bu hastalarda baş aşırı büyük, ekstremiteler ise oldukça kısadır. Gövde dardır ancak uzunluğu normaldir. Bu hastaların alnı belirgin, burun kökü basıktır. Yenidoğan refleksleri azalmıştır. Hidrosefali yaygındır. Doğumda inatçı bir siyanoz ve solunum solunum güçlüğü dikkati çeker. Olguların grafilerinde vertebra gövdesi basık, pedikülleri ise normaldir. Grafi bulguları; ön-arka grafide H veya U şeklinde vertebra, lateral grafide ise intervertebral aralıklar geniştir. İliak kemik dörtgeni andırır, asetabulum çatısı düzleşmiş, skapula ise daha küçüktür. Uzun kemikler kısa, diafizleri geniş, metafiz çizgisi düzensizdir. Uzun kemikler dörtgen şeklindedir (15).
Doğumdan önce, USG ile görülebi,len iskelet anomalilerine ek olarak, fetal hareket azlığı ve polihidroamniyoz gözlenmiştir. TD’nin FGFR3 mutasyonalrı ve radyolojik sınıflamalarına göre TD tip 1 ve TD tip 2 olmak üzere iki alt sınıfa ayrılmıştır (19).
2.4.4.1.1 Akondrogenezi I (Parenti-Fraccaro) ve Akondrogenezi II (Langer-Saldino)
Otozomal resesif geçişli olan iki sendromda da genellikle ölü doğum olur. Görülme sıklığı iki cinsiyette de eşittir. Bu hastalar büyük kafatası, oldukça kısa ekstremitelere, kısa ve geniş bir gövde yapısına sahiptir. Vertebra gövdesi ve sakrumun kemikleşmesi noksandır. Vertebra pedikülleri iyi gelişmemiştir. Bu
hastaların oldukça ince, kolay kırılgan kostaları vardır. Küçük ve kare şeklinde el ve ayakları vardır (15).
TD tip 1 olan hastalarda ciddi femur ve diğer tübüler kemiklerde eğrilme gözlenir. TD tip 1 genel olarak FGFR3 sistein rezidülerini oluşturan FGFR3 ekstraselüler domainin iki farklı bölgesinde missense mutasyon farklılaşması sonucu oluşur. Bu mutasyonlar genellikle ligand bağımsız reseptör dimerizasyonuna yol açar. RET protoonkojen ürünü, multiple endokrin neoplazi tip II-A’da görülen değişiklere benzer. Bazı değişikliklikler; stop kodonun arjinin ya da glisin ile değişimi sonucu 807 kodonda haritalanmıştır. Bu proteinin normalden 141 aminoasid daha uzun olması ile sonuçlanır (19).
Akondrogenezi II’de baş büyüklüğü ilk tipine göre daha orantılıdır. Kostalar kısadır. Kosta çapları ise normale yakındır (15).
Histolojik incelemede ‘’akondrogenezi’’ ismine tezat olarak; kıkırdak hücrelerinin volümlerinde artma, matriks yapısında azalma dikkat çekicidir. Kemikleşme çizgisi düzensizdir (15). TD2 genel olarak homojendir. FGFR3’ün distal intraselüler tirozin kinaz domainindeki 650. kodondaki Lys>Glu değişimi sonucu oluşur. FGFR3 aktivasyon alanına komşu glutamik asitteki mutasyon sonucu normal ligand bağımlı otofosforilasyonunu taklit eder. TD tip 2’li bireyler düz femurlara koronal ve lamboid suturların kranyosinostozuna (yonca yaprağı kafatası görünümü) sahiptirler (19).
2.4.4.2 Akondroplazi
İskelet displazilerinde en sık karşılaşılan gruptur (15). 1994 yılında, akondroplaziye neden olan fibroblast büyüme faktörü reseptörü 3 (FGFR3) geninde aktive edici mutasyonun tanımlanması , kondrodisplazilerin araştırma alanında önemli gelişmeye yol açmıştır. FGFR3’te diğer aktive edici mutasyonlar ise; akondroplazinin hafif bir klinik tablosu olan hipokondroplaziden, akondroplazinin oldukça ağır bir formu olan, gelişimsel gecikme, işitme kaybı, akantoz nigrikans ve ölümcül boy kısalığının eşlik ettiği geniş bir klinik tablo olan SADDAN’a yol açabilir (20).
Hastaların doğum sonrası muayenede farkedilebilecek kadar vücuda oranla ekstremite kısalığı belirgindir. Ortalama doğum boyu 47cm’dir. Hastalar brakisefaliktir ve kafatası büyülüğü >97 persentildir. Normal doğumda baş-pelvis