Molecular karyotyping of human hepatocellular carcinoma cell lines using single-nucleotide polymorphism arrays
Tam metin
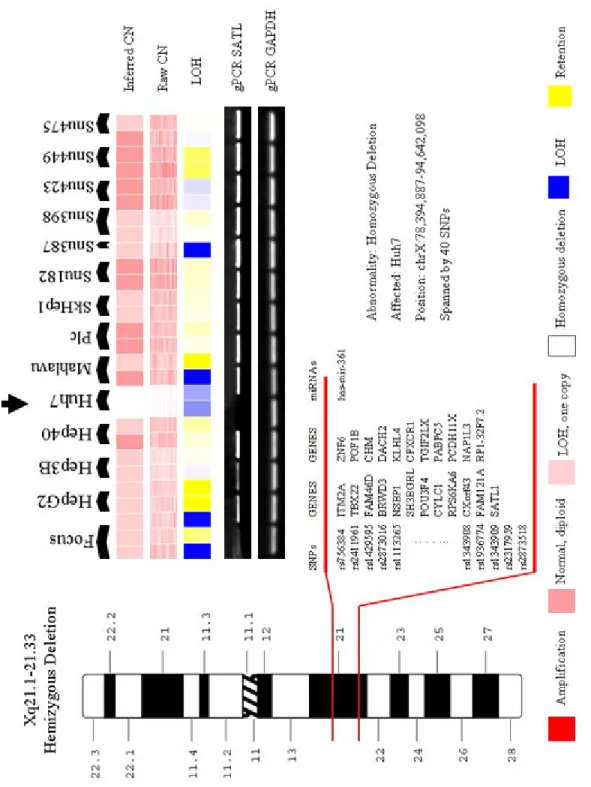
Şekil





Benzer Belgeler
雙和醫院 10 週年院慶,聚智創新邁向新北醫療生技城 雙和醫院在地深耕服務已邁入第 10 年,於 2018 年 6 月
— Mart 1945’den Haziran 1948’e kadar Eskişehir ve Adana’da yayınlanan Türke Doğru adlı derginin önce yönetmenliğini sonra yazı işleri müdürlüğünü
ue: upper epidermis; pp: palisade parenchyma; sp: spongy parenchyma; xy: xylem; le: lower epidermis; ph: phloem; sc:
It is interesting to note that since the sequential partitioner used during the initial partitioning phase is PaToH – a multilevel partitioner – the other parallel phases can be seen
antikor çeşitliliğini sağlamakta kullanılan sistemler aynıdır, aynı V(D)J rekombinaz kullanılır, ancak somatik hipermutasyon görülmez. uyarılmaları Antijen sunan
Figure 12.: Recombinant Activin-A was enhanced tumor growth. A) Relative luciferase activity of HepG2 cells 72h after treatment of Activin-A. B) Relative luciferase
The PROVEAN analysis identified D409H as accountable for harmful single nucleotide polymorphism, yet N370S for being a neutral amino acid substitution.Furthermore, L296V and
Anahtar Sözcükler: Parotis tümörleri, parotidektomi, nötrofil lenfosit oranı, platelet lenfosit oranı, sistemik inflamatuar index THE SIGNIFICANCE OF SYSTEMIC INFLAMMATORY
