Ecotype Traits of the Natural Populations of the
Birdsfoot Trefoil (
Lotus corniculatus) in Association
with the Geographical Parameters of the
Sampling Sites
Abstract
Birdsfoot trefoil, which is native to Turkish flora, is the most valuable and common Lotus species in Turkey. However, existing information on the ecology, distribution, and plant traits of the Birdsfoot trefoil is scant for the natural flora of Turkey. Therefore, seeds from 126 wild birdsfoot trefoil (WBT) populations were collected from plants spontaneously occurring in natural pasture and rangelands located (altitudes ranged from 5 to 2193 m a.s.l.) in the Black Sea Region of Turkey. These populations were grown under the same ecological conditions in 2010 and 2011 to determine the ecotype traits of the WBT and the relationships between these traits and the geographical features of the locations. The WBT was present (1-25% of botanical composition) in all ranges of altitude, although there was an increase in the presence and frequency of WBT with increasing altitude until 1000 m a.s.l. There were negative correlations between some traits (dry matter and seed yields, morphologic, and also root and crown-rot resistant traits) and altitude at which the seeds were collected. The data revealed that there was evidence of considerable inherent variations in all traits and thus there was a huge genetic diversity in the region. When populations were compared for agronomic and morphologic traits, the best populations were between altitudes of 251 and 500 m. It was observed that all populations had a spreading growth habit, except for 05TA01, 05TA02, and 60ER01 (semi-erect) and 60TU02 (semi-spreading). The results suggested that new grazing, forage types, and disease resistant WBT varieties may be bred from the studied populations.
Keywords: Birdsfoot trefoil, genetic resources, growth habit, stand persistence, variation
Örnekleme Sitelerinin Coğrafi Parametrelerine Bağlı Olarak Doğal Sarıçiçekli Gazal Boynuzu (Lotus corniculatus) Popülasyonlarının Ekotip Özellikleri
Özet
Sarıçiçekli gazal boynuzu (SGB), Türkiye’nin doğal florasında yer alan gazal boynuzu türleri içerisinde en değerlisi ve en fazla yayılım gösteren türüdür. Bununla birlikte bu türün Türkiye florasındaki ekolojisi, dağılımı ve bitkisel özellikleri hakkındaki bilgi oldukça sınırlıdır. Bu nedenle Türkiye’nin 5 ile 2193 m arasında değişen rakım değerlerine sahip Karadeniz Bölgesi doğal çayır ve meralarında kendiliğinden yetişen 126 SGB popülasyonu tohumları toplanmıştır. Bu popülasyonlar 2010 ve 2011 yılarında aynı ekolojik koşullarda yetiştirilerek bunların ekotip özellikleri ve bu özellikler ile toplama bölgelerinin coğrafi parametreleri arasındaki ilişkiler belirlenmiştir. İçinde bulundukları botanik kompozisyonların %1 ile 25’i arasında bir oran teşkil eden SGB bitkileri, çalışma alanının bütün rakım değerlerinde var olmasına karşın, bulunma sıklıkları deniz seviyesinden 1000 m rakım değerine kadar artış göstermiştir. İncelenen özelliklerden kuru madde ve tohum verimleri, morfoljik, kök ve köktacı hastalıklarına dayanım gibi özellikleri ile toplama yerlerinin rakım değerleri arasında negatif korelasyonlar belirlenmiştir. Araştırmadan elde edilen veriler, incelenen tüm özellikler bakımından popülasyonlar arasında dikkate değer kalıtsal varyasyonların olduğunu ve buna bağlı olarak çalışılan bölgede büyük bir genetik farklılık olduğunu göstermiştir. Agronomik ve morfolojik özellikler bakımından en iyi değerlere sahip olan popülasyonlar 251-500 m rakım aralığında yer almışlardır. Popülasyonların 122 adedi yatık, 3 adedi yarı dik ve 1 adedi ise yarı yatık gelişme göstermişlerdir. Bu sonuçlara göre, çalışılan popülasyonlardan otlatma ve yem tipi ile hastalıklara dayanaklı yeni çeşitler ıslah edilebilir.
Anahtar Kelimeler: Gazal boynuzu, genetik kaynaklar, gelişme formu, tesis ömrü, varyasyon.
Uzun F, Donmez HB (2016) Ecotype Traits of the Natural Populations of the Birdsfoot Trefoil (Lotus corniculatus) in Association with the Geographical Parameters of the Sampling Sites. Ekoloji 25(98): 33-40.
doi: 10.5053/ekoloji.2015.21
Received: 04.07.2014 / Accepted: 10.04.2015 Ferat UZUN1*, Hasan Beytullah DONMEZ2
1Department of the Field Crops, Faculty of Agriculture, Ondokuz Mayis University, 55139,
Samsun, TURKEY
2Organic Agricultural Programme, Cukurova University, Vocational School of Tufanbeyli,
01640, Adana, TURKEY
INTRODUCTION
The birdsfoot trefoil is primarily used for pasture but, it can also serve as hay and silage all over the world because of their plasticity and productivity in a wide range of soils and climatic conditions like other Lotus species (Blumenthal and McGraw 1999). The birdsfoot trefoil is considered one of the most important forage plants together with alfalfa and clover. On the other hand, during the last few years the birdsfoot trefoil has gained in popularity due to three main comparative advantages. Firstly, the interest in low input systems revalidates the place of birdsfoot trefoil as an economic species with low resource requirements. Secondly, the birdsfoot trefoil feeding value has particular advantages over many other legume species due to the presence of condensed tannins (Waghorn 2008). Thirdly, a new achievement has been obtained for greater persistence against crown rot pathogens (Beuselinck et al. 2005).
The birdsfoot trefoil is an excellent resource to improve pasture and rangelands in Turkey as well as in most parts of the world. Although determination of variation for quantitative and qualitative traits of the plants genetic resources growing in natural pasture and rangeland habitats is important for breeding programmes, scientific researches on the wild birdsfoot trefoil, including its ecology and distribution in Turkey is quite low (Uzun et al. 2008). Therefore, the objectives of this research was to determine these traits of some of the wild birdsfoot trefoil populations sampled from the Black Sea Region of Turkey, and also to determine the relationships of these traits with the geographic features of the sampling sites.
MATERIALS AND METHODS
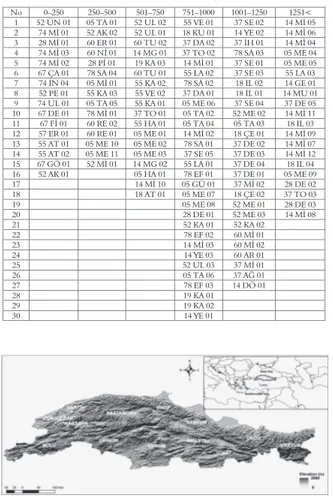
A total of 126 wild populations of birdsfoot trefoil seeds were collected from the natural areas of the Turkish Black Sea Region (31°33’- 37°32’ E and 40°06’ - 42°03’ N), in the summer of 2009 and 2010. The investigation area has an irregular topography with elevations from 5 to about 2193 m (Table 1 and
sampling site was nearly 10 acres. In each population, the mature pods were randomly sampled on a site with uniform growing conditions. Each population was represented by a minimum of 50 plants (Brown and Marshall 1995). Five ranges of altitude were adopted for the classification of wild birdsfoot trefoil populations (Carneiro and Serrao 1996).
The glasshouse and field studies of this study were carried out in the Black Sea Agricultural Research Institute, Samsun, between the years 2010 and 2012. The experimental site was located at 41°17’ N, 36°21’ E, at an elevation of 4 m. The soil was 2.40% clay and a pH of 7.64. The area has a mild and humid climate with a mean annual temperature of 14.4°C and a mean annual precipitation of 675.1 mm (1974-2013 years) (Anonymous 2013).
The seeds of each population were sown into plug trays containing peat and grown in glasshouse conditions under natural light. Forty-five seedlings from each population were randomly selected and transplanted into the field with three replications at the 8-10 cm length stage at spacing of 0.9 х 0.9 m at the end of November 2010.
To determine the dry matter (DM) yield, the 15 plants from each population were harvested manually three times in 2011 and two times in 2012 at the beginning of the flowering stage. When 65 -70% of the pods turned brown in colour, the other 15 plants were harvested for seed yield. The remaining 15 plants were used to determine the morphologic and phenologic traits. These traits were measured and evaluated according to Anonymous (1991).
The stand persistence ratios of the populations were determined at the end of the first and second growing season. In this study, all data regarding the studied traits were obtained in the first experimental year. In addition, the DM and seed yields, and disease severity were determined in the years 2011 and 2012.
RESULTS AND DISCUSSION Distribution of Populations Regarding Altitude
The distributions of the birdsfoot trefoil populations based on the altitude of the collecting sites are given in Table 1. The numbers of the collected birdsfoot trefoil populations were 13% at 0-250 m, 12% at 251-500 m, 14% at 501-750 m, 24% at 751-1000 m, 21% at 1001-1250 m, and 16% at 1251-2193 m. These results indicate that this species may be adapted to all ranges of altitude, as reported by Davis (1970) and Sönmez (2005). Chrtkova-Zertova (1973) also stated that vertical distribution of birdsfoot trefoil extends from the coastal zone to the alpine zone at all altitudes above sea level. This is consistent with the idea that the ecological range of the Lotus species is very wide, corresponding to its polymorphous character (Drobna 2010). On the other hand, there was an increase in the present
frequency of birdsfoot trefoil increasing with altitude till 1000 m. Based on the findings of the present study and some other studies (Steiner and Santos 2001, Uysal et al. 2011), it can be said that birdsfoot trefoil is better adapted to the high-elevation areas.
Description of Populations
There were considerable variations for the morphologic, agronomic, and phenotypic traits of the populations, as stated by Steiner and Santos (2001).
Leaves were characterized as having three leaflets per leaf and two leaf-like stipules at the base of each leaf for all populations. The populations were variable for the middle leaflet length and width. The middle leaflets size ranged between 0.65-1.86 in length and 0.30-0.90 cm in width (Table 2) with the 05TA01 population having the longest and also widest leaves.
The populations differed in the number of stems per plant, which ranged from 12.17 to 51.33. The highest number of stems per plant was found in 74IN04, 78SA04, and 18CE02 and the lowest was in the 52KA02 population. The length of the main stem varied among the populations was between 18.00 (28DE03) and 80.22 (05ME11) cm. The populations were also quite variable for stem height. The stem height of the populations were between 3.42 and 54.32 cm (Table 1). The highest stem heights were measured in 05TA02, 60ER01, and 05ME10, and the lowest in the 14Mİ08 population (Table 2).
In this study, it was observed that the populations had a mostly spreading growth habit, except for four populations. The populations 05TA01, 05TA02, and 60ER01 had semi-erect habit, while 60TU02 had a semi-spreading growth habit and there were no rhizome.
As known, shorter forage types are considered more suitable for grazing than taller ones. Indeed, sampling sites of the populations have been grazed directly by livestock throughout year. Thus, new varieties for grazing types can be selected from these decumbent populations and were determined to be a highly variable. On the other hand, selection studies can be conducted for hay types from semi erect populations. Semi-erect and semi-spreading populations also have spreading stems. Therefore, from these populations new grazing type varieties can also be selected.
Table 1. Distribution of 126 wild birdsfoot trefoil populations based on the altitude of the sampling sites (m) in the Black Sea Region of Turkey.
Fig. 1. Study area from where the birdsfoot trefoil populations were sampled.
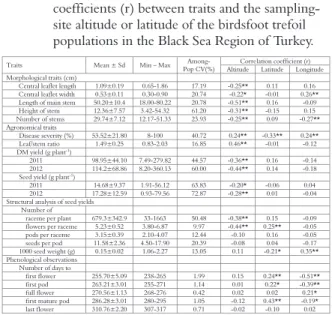
The number of raceme per plant, number of flowers, and pods per raceme varied from 33 (28DE03) to 1663 (05TA01), 3.80 (52UL02 and 14MI08) to 6.87 (55VE02), and 2.10 (05GU01) to 4.07 (60RE02), respectively. The maximum number of seeds per pod was obtained in 52KA01 (17.90), and the minimum in the 28DE03 (4.50) population. There were significant correlations between the leaflet length (r=-0.25*), leaflet width (r=-0.22*), stem number 0.25**), stem length (r=-0.51**), stem height (r=-0.31**), number of raceme per plant (r=-0.38**) or number of flowers per raceme (r=-0.43**), and altitude. In addition to, the correlations between the number of flowers per raceme (r=0.25**) and the latitude; leaflet width (r=0.26**) or number of stems per plant (r=-0.27**) and longitude were statistically significant (Table 2).
As shown in Table 2, the average DM yields were 98.95 and 114.20 g plant-1in the first and the second
years, respectively. The population 05TA01 had the highest mean DM yields per plant (279.82 g plant-1),
diet selection, and forage intake (Forbes and Coleman 1993). This trait has been used successfully to predict the forage quality parameters in the birdsfoot trefoil (Buxton et al. 1985). Also, these authors reported that attempts to improve the forage quality of birdsfoot trefoil should be directed toward the lower portion of the plant canopy and stem. Based on observation in this study, it can be said that the hay quality of the 37MI01, 37MI02, and 52UN03 populations were higher than others due to the high leaf to stem ratio.
The populations were also quite variable for seed yield. The average seed yields were 14.68 and 17.28 g plant-1in 2011 and 2012, respectively. The reason
of the differences of seed yields in first and second year might be attributed to the differences of the insect pollinator amount and activity in these years due to different climatic conditions. The highest seed yields per plant were 56.12 and 79.56 g plant-1
in population of 05TA02 in the first and second year, respectively. The population of 14MI08 (1.91 g plant-1) had the lowest seed yield in the first year and
the 14MI08 (0.95 g plant-1) and 52KA02 (0.93 g
plant-1) in the second year.
The 1000 seed weigh ranged between 1.06 (74IN01) and 2.27 g (28DE01). It is showed that the 1000 seed weight is under strong genetic control.
There were significant correlations between the DM yield (0.36** in the first year and r=-0.44** in the second year), leaf/stem ratio (r=0.43**), seed yield (0.20* in 2011 and r=-0.28** in 2012), and disease severity (0.24**) and altitude. Furthermore, latitude and longitude were highly correlated with the 1000 seed weight (r=-0.21* and 0.35**, respectively) and disease severity (r=-0.33** and r=0.24**, respectively).
The first flower, full flower, and end of the flowering stage of the populations ranged between 238 and 265, 268 and 276, and 307 and 317 days, respectively. The mean number of days to first pod and mature pod varied from 255 to 271 and from 280 to 295 days, respectively. No significant
Table 2. Descriptive statistics of the morphological, agronomical, phenological traits, and structural analysis of the seed yields and also the correlation coefficients (r) between traits and the sampling-site altitude or latitude of the birdsfoot trefoil populations in the Black Sea Region of Turkey.
and first mature pod (r=-0.19*).
The results related to the morphological traits were corroborated with those obtained by Drobná (2010), who reported that the most variable were the quantitative traits related to the stem and number of inflorescence per stem. Variability also exists for specific agronomic traits, including herbage yield and quality, flowering habit, and reproductive compatibility (Santos et al. 2001, Steiner and Santos 2001). The morphological traits of plants in terms of size and growth habit are influenced by the genotype and environment or their interaction. Furthermore, morphological variations among populations are strongly associated with the geographical features of the collecting-site locations. In the present study, considerable variations for the morphological traits of populations in terms of size and reproductive growth resulted in the genotype due to the populations grown in the same environment. The number of stems per plant (r=0.44**), stem length (r=0.68**), and stem height (r=0.66**) were the main contributor to the DM yield (data not shown in table) because the general view of the plants ranged from small and weak to large plants with a high number of stems (Vuckovic et al. 2007, Churkova 2011). Our results on the variation in the morphological traits support the idea that variation in morphological traits are revealed by the existence of considerable variation in the form and growth of the plants such as stem length and shape, number of flowers in inflorescence etc. The high variability in these morphological traits might be related to the physiological adaptation of the plants to their environments (Steiner and Santos 2001), because of the strong relationship among the morphological traits of birdsfoot trefoil and the geographical features of the sampling-sites. The morphological similarities among the genotypes may be related to the similarities of the general ecogeographic features of the sampling sites as reported by Steiner and Santos (2001).
The morphological and agronomical variations were strongly associated with the altitude of the sampling-site. The leaflet, stem, and raceme traits, and the DM and seed yields and leaf to stem ratio were influenced highly by the altitude (Table 2). All morphological traits exhibited negative correlation with altitude in our study. Most of the populations from locations from a lower altitude tended to
produce plants with high DM yield, number of longer stems, seed yield and the number of raceme per plant, inflorescence per plant and seed per pod etc. compared to populations from locations from higher altitudes as stated by Drobna (2010). Stoffella et al. (1998) noted that birdsfoot trefoil ecotypes can be distinguished by morphological attributes. The birdsfoot trefoil populations studied in the present study were distinguished by morphological attributes using the standard division of the mean. This analysis shows that the within-population variation was larger than the among-population variation for most evaluated traits (data not shown), as reported by Julier et al. (2000) and Drobna (2010) for birdsfoot trefoil and alfalfa, which are strongly cross-pollinated species. Assuming that some of these specific variations have a genetic basis, this may suggest that there is a potential for improving the birdsfoot trefoil which might allow the populations to be productive and persistent.
A major factor limiting more widespread use of the Lotus species is the fact that achieving a high seed yield in these species is difficult (McGraw and Beuselinck 1983). In high variation in terms of seed yield of the Lotus species, the number of raceme per plant, number of pod per raceme, number of seed per pod, and seed weight have been identified as major contributing traits (Stephenson 1984, McGraw et al. 1986, Garcia-Diaz and Steiner 2000). Furthermore, Li and Hill (1989) and Qingfeng (1989) reported that the seed yield is primarily determined by the number of inflorescences produced per plant, in turn, the inflorescence number is primarily dependent on the number of shoots available at the time of flowering. In addition to these factors the insect pollination amount and activity is also highly effective for the legume species seed yield. In the present study, there are the significant positive relationships between seed yield and leaflet length (0.42**), leaflet width (0.33**), stem number (0.27**), stem length (0.45**) stem height (0.62**), raceme number per plant (0.58**), flower number per raceme (0.39**), herbage yield (0.57**), number of days to first flower (0.24**), first pod (0.30**), and full flower (0.18*). Therefore, it can be suggested that these traits should be used as a selection criteria for improving seed production in the Lotus species.
A high phenotypic variability among the populations in terms of studied phenological traits
can be related to a high phenotypic plasticity and/or genetic variations among the individuals (Ferraro et al. 2010). This causes the spreading of birdsfoot trefoil in the heterogeneous environment. Indeed, genetic differentiation in phenotypic traits has been frequently demonstrated among the populations of wide-ranging species (Quinn and Wetherington 2002). The flowering habit of birdsfoot trefoil is indeterminate, so flowering is extended over a long period of time. Qingfeng (1989) reported that the protracted flowering pattern in birdsfoot trefoil is caused by the continuous production of shoots with the capacity to flower. Therefore, the continuous development of new shoots replacing older flowering shoots was the main cause of the long flowering period. On the other hand, the long flowering periods of populations may be advantageous for honey production.
Stand Persistence of Populations
There was no stand mortality at the end of the first year. There was an important reduction in the surviving plants and the stand mortality ratio of the populations changed from 8 to 100% at the end of the second year. The 14DÖ01 and 55KA03 were the most resistant populations for rot and root-crown diseases.
Lotus species have some advantages, such as
adaptation capability and high nutritive value and non-bloating compared to other many forage legumes, but it has poor stand persistence (Altier and Kinkel 2005). This author reported stand losses of 68 to 90% within 2 year of establishment due to root and crown-rot diseases for birdsfoot trefoil. The results of our study were similar to the study of these authors. Formoso (1993) also stated that significant plant losses were observed in pastures two years old and older in Uruguay, especially following a period of summer drought which resembles our experimental area. Breeding for disease resistance generally has involved finding suitable genetic material amongst existing stocks or in the wild. Astley (1987) reported that germplasm
improve stand persistence (Beuselinck et al. 2005). However, no rhizomatous growth form within the populations was observed.
Principle Component and Cluster Analysis
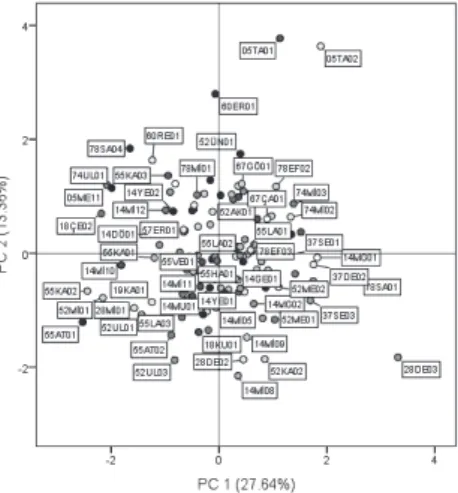
To assess the relative significance of evaluated traits, a PCA was carried out. The PCA revealed that the first six PCs explained 70.06% of the total variation. The attributes that appeared to be the major sources of diversity within the evaluated populations were the leaflet length, length of main stem, height of main stem, number of raceme per plant, number of flowers per raceme, dry matter, and seed yields (PC1). The number of days to first flower, first pod, full flower, and first mature pod are explained in PC2. The leaflet width are explained in PC3. The 1000 seed weight and number of days to last flower are explained with PC4 and the number of pods per raceme are explained in PC5. Therefore, the above-mentioned traits must be primarily taken into consideration for selection.
Based on the distance matrix for the distributions of the populations on the first two axes, some populations (e.g. populations of 28DE02, 14MI08, and 52KA02 etc.) were closely similar to each other while some (e.g. populations of 18CE02, 05TA01, and 28DE03 etc.) were notably quite
semi spreading and also have spreading stems, from these populations selections can be made for new hay and grazing type varities for improving pasture and rangeland where alfalfa and other legumes cannot grow well due to infertile, acidic, drought, or wet soil conditions. The 05TA02 is also the most promising population for seed yield. On the other hand, 14DÖ01 and 55KA03 were the most resistant populations for rot and root-crown diseases, therefore, these populations can be very important
in resistance studies in the future. When populations were compared for DM yield, plant height and length, stem number, and disease resistance, the best populations were found between the 250 and 500 m elevations.
ACKNOWLEDGEMENTS
This research was supported by The Scientific and Technological Research Council of Turkey (Project No. TOVAG 108O658).
REFERENCES
Altier NA, Kinkel LL (2005) Epidemiological studies on crown and root rot of birdsfoot trefoil in Uruguay. Lotus Newsletter 35(1): 42-58.
Anonymous (1991) Descriptor list for forage legumes (Birdsfoot trefoil). International Board for Plant Genetic Resources, Rome.
Anonymous (2013) Turkish State Meteorological Service Records of Samsun Province, Samsun. Astley D (1987) Genetic resource conservation. Experimental Agriculture 23: 245-257.
Beuselinck PR, Steiner JJ (1992) A proposed framework for identifying, quantifying, and utilizing plant germplasm resources. Field Crops 29: 261-272.
Beuselinck PR, Brummer EC, Viands DK, Asay KH, Smith RR, Steiner JJ, Brauer DK (2005) Genotype and environment affect rhizome growth of birdsfoot trefoil. Crop Science 45: 1736-1740.
Blumenthal MJ, McGraw RL (1999) Lotus Adaptation, Use and Management. In: Beuselinck PR (ed), Trefoil: The science and Technology of Lotus, American Society of Agronomy, Madison, 97-119.
Brown AHD, Marshall DR (1995) A Basic Sampling Strategy: Theory and Practice. In: Guarino L, Ramanatha RV, Read R (eds), In Collecting Plant Genetic Diversity: Technical Guidelines, Wallingford, 75-92. Buxton DR, Hornstein JS, Wedin WF, Marten GC (1985) Forage quality in stratified canopies of Alfalfa, birdsfoot trefoil and Red Clover. Crop Science 25: 273-279.
Carneiro JP, Serrao MG (1996) Programs for improvement of Medicago sp. at ENMP-Portugal. I. distribution of annual medics as related to soil properties and climatic conditions. CIHEAM Options Mediterraneennes 18: 23-29.
Chrtkova-Zertova A (1973) A monographic study of Lotus corniculatus L. I central and northern Europe. Rozpravy Ceskoslovenske Akademie Ved 83: 1-94.
Churkova B (2011) Correlations dependence and degree of variation between yield and some morphological parameters in birdsfoot trefoil (Lotus corniculatus L.) accessions. Bulgarian Journal of Agricultural Sciences 17(4): 37-441.
Davis PH (1970) Flora of Turkey and East Aegean Islands. Edinburg University Press, Edinburg.
Drobna J (2010) Morphological variation in natural populations of Lotus Corniculatus in association to geographical parameters of collecting sites. Biologia, Section Botany 65(2): 213-218.
Ferraro G, Filip R, Pero M, Basualdo N, Mendoza R, Garcia I (2010) Flavonoids of Lotus tenuis (Waldst.&Kit.) as markers of populations growing in soils of different saline and hydrologic conditions. Journal of the Brazilian Chemical Society 21(9): 1739-1745.
Forbes TDA, Coleman SW (1993) Forage intake and ingestive behaviour of Cattle grazing old world bluestems. Agronomy Journal 85: 808-816.
Formoso F (1993) Lotus corniculatus. I. Performance Forrajera y Caracteristicas Agronomicas Asociadas. INIA Serie Tecnica, Montevideo.
Garcia-Diaz CC, Steiner JJ (2000) Birdsfoot trefoil seed production: II. Plant water status on reproductive development and seed yield. Crop Science 40: 449-456.
Julier B, Huyghe C, Ecalle C (2000) Within and among cultivar Genetic variation in Alfalfa: Forage quality, morphology and yield. Crop Science 40: 365-369.
Li Q, Hill ML (1989) A study of flower development and seed yield components in birdsfoot trefoil (Lotus
corniculatus L.). Journal of Applied Seed Production 7: 65-69.
McGraw RL, Beuselinck PR (1983) Growth and seed yield characteristics of birdsfoot trefoil. Agronomy Journal 75: 443-446.
McGraw RL, Beuselinck PR, Ingram KT (1986) Plant population density Effects on seed yield of birdsfoot trefoil. Agronomy Journal 78: 201-205.
O'Rourke N, Hatcher L, Stepanski EJ (2005) A Step-by-step Approach to Using SAS for Univariate and Multivariate Statistics. Wiley, New York.
Qingfeng L (1989) Seed production in birdsfoot trefoil (Lotus corniculatus L.). PhD Thesis, University of Massey, Palmerston.
Quinn JA, Wetherington JD (2002) Genetic variability and phenotypic plasticity in flowering phenology in populations of two grasses. Journal of the Torrey Botanical Society 129(2): 96-106.
Santos G, Steiner JJ, Beuselinck PR (2001) Adaptive ecology of Lotus corniculatus L. genotypes: II. Crossing ability. Crop Science 41: 564-570.
Sönmez S (2005) Kurtçalı Tepe (Balıkesir) ağaçlandırma sahasında vejetasyonun süksesyonel değişimi ve floristik gözlemler. Ekoloji 14(57): 1-12.
Steiner JJ, Santos GG (2001) Adaptive ecology of Lotus corniculatus L. genotypes. I. Plant morphology and RAPD Marker Characterizations. Crop Science 41: 552-563.
Stephenson AG (1984) The regulation of maternal investment in an indeterminate flowering plant (Lotus
corniculatus L.). Ecology 65(1): 113-121.
Stoffella SL, Posse G, Collantes M (1998) Phenotypic plasticity and genetic variation of Lotus tenuis populations from soils with different pH. Ecologia 8: 57-63.
Uysal I, Karabacak E, Oner S, Kurt F (2011) A syntaxonomical study of the pseudo-alpine vegetation of Kazdagi and two new endemic associations. Ekoloji 20(80): 88-96.
Uzun F, Sulak M, Ugur S (2008) The important of birdsfoot trefoil species for Turkey. Turkish Journal of Scientific Reviews 1(2): 45-54.
Vuckovic S, Stojanovic I, Prodanovic S, Cupina B, Ivanovic T, Vojin S, Jelacic S (2007) Morphological and nutritional properties of birdsfoot trefoil (Lotus corniculatus L.) autochthonous populations in Serbia and Bosnia and Herzegovina. Genetic Resources and Crop Evolution 54(2): 421-428.
Waghorn G (2008) Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production-progress and challenges. Animal Feed Science and Technology 147: 116-139.