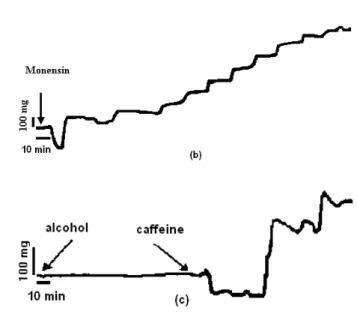
Investigation of the cellular mechanisms of actions of digoxin and monensin by using guinea-pig papillary muscles
Tam metin
Şekil

Benzer Belgeler
Research Author(s) Research Design Employed Techniques FL Has Positive Effect on Sohrabi and Mohammadi, 2019; Mixed Research Design Quick placement test, observations,
In our case, we think that elevation in the central venous pressure accompanied by left and right heart failure, induced by nonischemic heart disease, might have increased
In this preliminary study, unstable patient presenting with dyspnea, we investigated the diagnostic value of s-trem to differentiate whether the etiology is sepsis- induced
Chapter 2 looked at the disallowance of violence through arguments of morality and efficacy, and argued that nonviolence assumes the legitimacy of the order and the state;
According to the anatomical compatibility of the right renal artery and after consultation with the vascular surgery department, percutane- ous closure with Amplatzer vascular
Organization of heart failure management in European Society of Cardiology member countries: survey of the Heart Failure Association of the European Society of Cardiology in
The other calculated parameters were as follows: corrected serum sodium, 120.7 mmol/L; measured serum osmolality, 276 mOsm/kg (reference range 275–295 mOsm/kg); and effective
In the Acute and Chronic Heart Failure Diagnosis and Treatment Guidelines published by the ESC in 2012, ARBs are recommended as class 1, evidence level A for HFrEF patients not
