T.C.
SELÇUK ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
DEĞİŞİK DOZLARDA UYGULANAN AZOTUN BAZI EKMEKLİK BUĞDAY
ÇEŞİTLERİNİN KARBONHİDRAT TRANSLOKASYONUNA ETKİSİNİN
BELİRLENMESİ Oğuz ÖNDER DOKTORA TEZİ
Toprak Bilimi ve Bitki Besleme Anabilim Dalı
Ocak-2015 KONYA Her Hakkı Saklıdır
TEZ KABUL VE ONAYI
Oğuz Önder tarafından hazırlanan “Değişik Dozlarda Uygulanan Azotun Bazı Ekmeklik Buğday Çeşitlerinin Karbonhidrat Translokasyonuna Etkisinin Belirlenmesi” adlı tez çalışması 05/01/2015 tarihinde aşağıdaki jüri tarafından oy birliği ile Selçuk Üniversitesi Fen Bilimleri Enstitüsü Toprak Bilimi ve Bitki Besleme Anabilim Dalı’nda DOKTORA TEZİ olarak kabul edilmiştir.
Jüri Üyeleri İmza
Başkan
Prof.Dr. Ali TOPAL ………..
Danışman
Prof.Dr. Sait GEZGİN ………..
Üye
Prof.Dr. Ali TOPAL ………..
Üye
Prof.Dr. Bülent TORUN ………..
Üye
Doç.Dr. Mehmet ZENGİN ………..
Üye
Doç.Dr. Mehmet HAMURCU ………..
Yukarıdaki sonucu onaylarım.
Prof. Dr. Aşır GENÇ FBE Müdürü
Bu tez çalışmasının bir kısmı Selçuk Üniversitesi, Bilimsel Araştırma Projeleri Koordinatörlüğü tarafından 11201088 nolu proje ile bir kısmı da TAGEM/TBAD/12/A12/P01/07-001 tarafından desteklenmiştir.
TEZ BİLDİRİMİ
Bu tezdeki bütün bilgilerin etik davranış ve akademik kurallar çerçevesinde elde edildiğini ve tez yazım kurallarına uygun olarak hazırlanan bu çalışmada bana ait olmayan her türlü ifade ve bilginin kaynağına eksiksiz atıf yapıldığını bildiririm.
DECLARATION PAGE
I hereby declare that all information in this document has been obtained and presented in accordance with academic rules and ethical conduct. I also declare that, as required by these rules and conduct, I have fully cited and referenced all material and results that are not original to this work.
Oğuz ÖNDER 05/01/2015
iv
ÖZET
DOKTORA TEZİ
DEĞİŞİK DOZLARDA UYGULANAN AZOTUN BAZI EKMEKLİK BUĞDAY ÇEŞİTLERİNİN KARBONHİDRAT TRANSLOKASYONUNA ETKİSİNİN
BELİRLENMESİ
Oğuz ÖNDER
Selçuk Üniversitesi Fen Bilimleri Enstitüsü Toprak Bilimi ve Bitki Besleme Anabilim Dalı
Danışman: Prof.Dr. Sait GEZGİN 2015, 275 Sayfa
Jüri
Prof.Dr. Sait GEZGİN Prof.Dr. Ali TOPAL Prof.Dr. Bülent TORUN Doç.Dr. Mehmet ZENGİN Doç.Dr. Mehmet HAMURCU
Sapta biriktirilen karbonhidratlar tane doldurmada önemli bir karbon kaynağıdır. Biyolojik verimin artırılması, çevresel ve biyotik stres faktörlerine karşı, tane doldurmayı destekleyen özelliği nedeniyle ıslahta önemlidir. Buğday genotipleri arasında kuraklığa mukavemeti yüksek olanların belirlenmesinde kullanılacak parametrelerin geliştirilmesine ihtiyaç vardır. Bu amaçla Eskişehir koşullarında Bezostaja 1, ES 26, İzgi 2001 ve Müfitbey buğday çeşitleri, 0, 5, 10, 15 kg N/da olmak üzere dört farklı azot dozunda kuru (yağmura bağımlı) ve sulu (takviye sulamanın yapıldığı) şartlarda 2011 ve 2012 yıllarında yetiştirilmiştir. Araştırma tesadüf bloklarında, bölünmüş parseller deneme deseninde 4 tekerrürlü olarak yürütülmüştür.
Bu çalışmada; verim komponentleri, başaklanma tarihi, bitki boyu, bayrak yaprak alanı, biyolojik verim, gibi parametrelerin yanı sıra, fizyolojik parametrelerden; suda çözünebilir karbonhidrat içeriği (SÇK), translokasyon oranı, kurak hassasiyet indeksi (KHİ), kuru madde translokasyonu ve azot tekrar taşıma (ATT) ölçüm ve gözlemleri yapılarak, bu parametreler arasındaki ilişkiler araştırılmıştır.
Araştırma sonucunda; tane doldurmaya katkısı yönünden geççi çeşitlerde translokasyonla taşınan fotoasimilant miktarı ve ATT’nın erkencilere göre daha önemli olduğu görülürken, SÇK’ların özellikle kurak koşullarda translokasyonu desteklediği anlaşılmıştır. Çiçeklenme döneminde sapta bulunan SÇK’ların özellikle kuru koşullarda tane verimine olumlu katkı yaptıkları, ikinci (penultimate) boğumdan yapılacak SÇK ölçümlerinin genotipler arası farklılığı daha iyi ortaya koyduğu ve artan SÇK miktarının KHİ’ni düşürerek, verim stabilitesine olumlu katkı yaptığı belirlenmiştir.
v
ABSTRACT
Ph.D THESIS
DETERMINATION OF EFFECTS OF DIFFERENT NITROGEN DOSES ON CARBOHYDRATE TRANSLOCATION OF SOME BREAD WHEAT
VARIETIES
Oğuz ÖNDER
THE GRADUATE SCHOOL OF NATURAL AND APPLIED SCIENCE OF SELÇUK UNIVERSITY
THE DEGREE OF DOCTOR OF PHILOSOPHY IN SOIL SCIENCE AND PLANT FEEDING
Advisor: Prof.Dr. Sait GEZGİN 2015, 275 Pages
Jury
Prof.Dr. Sait GEZGİN Prof.Dr. Ali TOPAL Prof.Dr. Bülent TORUN Doç.Dr. Mehmet ZENGİN Doç.Dr. Mehmet HAMURCU
Water-soluble carbohydrates accumulated in stem are an important carbon sources for grain filling. Development of biomass, against abiotic and biotic stress factors, it is important in breeding, because of supporting features to grain filling. Among wheat genotypes which are high drought resistance to be used in determining, there is need for the development of parameters.
For this purpose; In Eskisehir conditions Bezostaja 1, ES 26, İzgi 2001 and Müfitbey wheat varieties, four different nitrogen (0, 5, 10, 15 kg N/da) application in dry (rainfed) and irrigated (reinforcement irrigation) conditions has grown year 2011 and 2012. The experiments carried out in randomized complete block, split plots in four replications.
In this study; yield components, heading date, plant height, leaf area, biomass such as parameters, as well as physiological parameters; water-soluble carbohydrate (WSC), translocation rate, drought sensitivity index (DSI), dry matter translocation and nitrogen remobilization (NR) measurements observed and between this parameters relationships were investigated.
As a result; in terms of its contribution to grain filling, in late heading varieties, amount of carried photoassimilant with translocation and NR is more important than early, while WSC, especially in rainfed conditions determined to support translocation. In flowering stage available WSC in stem especially rainfed conditions, makes positive contribution to grain yield, using second (penultimate) node to WSC measurements, better revealed differences between genotypes, and increasing WSC content reducing DSI was determined to make positive contribution to yield stability.
vi
ÖNSÖZ
Doktora çalışmalarımda, gerek derslerimde ve gerekse tez çalışmalarında, bana danışmanlık ederek, beni yönlendiren ve her türlü olanağı sağlayan danışmanım Sayın Prof.Dr. Sait GEZGİN’e;
Derslerimde ve tez çalışmalarımda değerli katkılar sağlayan, Sayın Prof.Dr. Ali TOPAL’a ve Sayın Doç.Dr. Mehmet HAMURCU’ya;
Araştırmanın başlangıcından sonuna kadar teorik ve pratik bilgilerinden yararlandığım Sayın Müfit KALAYCI’ya, Dr. Cemal ÇEKİÇ’e, Dr. Erdinç SAVAŞLI’ya, Dr. Fatma GÖKMEN’e, Zir.Müh. Ramis DAYIOĞLU’na, Zir.Müh. Burçin KISA’ya, Tek. Nuri TUNCER’e ve Tek. Mümtaz EKİZ’e;
Çalışmaların gerçekleştirilmesindeki her türlü desteklerinden dolayı Selçuk Üniversitesi’ne, Tarımsal Araştırmalar ve Politikalar Genel Müdürlüğü’ne ve Geçit Kuşağı Tarımsal Araştırma Enstitüsü’ne;
Eşim Hatice Selcan, oğlum Emre’ye, Ailem, Anne ve Baba’ma, En içten teşekkürlerimi sunarım.
Oğuz ÖNDER KONYA-2015
vii İÇİNDEKİLER ÖZET ... iv ABSTRACT ...v ÖNSÖZ ... vi İÇİNDEKİLER ... vii SİMGELER VE KISALTMALAR ... ix 1. GİRİŞ ...1 2. KAYNAK ARAŞTIRMASI ...3 3. MATERYAL VE YÖNTEM ... 12 3.1. Materyal ... 12 3.2. Deneme Deseni ... 13
3.3. Deneme Yerinin Meteorolojik Özellikleri ... 13
3.4. Tarla Denemeleri Ekim ve Hasat Zamanları ... 15
3.5. İstatistiki Analiz ... 17
3.6. Denemelerde Alınan Ölçüm ve Gözlemler ... 17
3.6.1. Translokasyon kapasitesi ... 17
3.6.2. Sapta, suda çözünebilir karbonhidratlar ... 18
3.6.3. Kuraklık hassasiyet indeksi ... 19
3.6.4. Bayrak yaprak yeşil kalma süresi ... 19
3.6.5. Yaprak alan indeksi ... 20
3.6.6. Biyokütle ağırlığı ... 20
3.6.7. Tozlanma öncesi asimilantların tekrar taşınması (remobilization) ... 22
3.6.8. Bitki boyu ... 23
3.6.9. Üst boğum (peduncle) uzunluğu ... 23
3.6.10. Sap doluluk oranı ... 23
3.6.11. Başaklanma-olum süresi... 24
3.6.12. Tane verimi ... 24
4. ARAŞTIRMA SONUÇLARI VE TARTIŞMA ... 25
4.1. Translokasyon Kapasitesi ... 25
4.2. Sapta, Suda Çözünebilir Karbonhidratlar ... 29
4.2.1. Suda Çözünebilir Karbonhidrat oranı ... 30
4.2.2. Kuru ağırlık esasına göre toplam SÇK miktarları ... 51
4.3. Kurak Hassasiyet İndeksi ... 85
4.4. Bayrak Yaprak Yeşil Kalma Süresi ... 87
4.5. Yaprak Alan İndeksi ... 91
4.6. Biyolojik Verim ve Kardeş Sayıları ... 97
4.6.1. Biyolojik verim ... 97
4.6.2. Metrekarade kardeş (sap) sayısı ... 118
viii
4.7.1. Kuru madde translokasyonu (KMT) ... 135
4.7.2. Kuru madde translokasyon etkinliği (KMTE) ... 141
4.7.3. Tozlanma öncesi asimilatların taneye katkısı (TÖAK) ... 145
4.7.4. Başak hasat indeksi ... 149
4.7.5. Erken dönem başak indeksi ... 155
4.7.6. Bitkide ve tanede azot kapsamı ... 160
4.7.7. Bitkide ve tanede azot miktarı ... 173
4.7.8. Azot tekrar taşıma (ATT) ... 183
4.7.9. Azot tekrar taşıma etkinliği (ATTE) ... 188
4.7.10. Tanede azot kapsamı üzerine, azot tekrar taşınmasının katkısı (ATTK) .. 194
4.7.11. Azot hasat indeksi ... 198
4.8. Bitki Boyu ... 201
4.9. Birinci Boğum (peduncle) Uzunluğu ... 207
4.10. İkinci Boğum (penultimate) Uzunluğu ... 212
4.11. Bayrak Yaprak Alanı ... 214
4.12. Sap Doluluk Oranı ... 216
4.12.1. Sapta özle dolu alan ... 216
4.12.2. Sapta özle dolu hacim ... 220
4.13. Başaklanma Olum Süresi ... 227
4.14. Tane Verimi ve Verim Unsurları ... 232
4.14.1. Tane verimi ... 232
4.14.2. Hasat indeksi ... 236
4.14.3. Metrekarede tane sayısı ... 240
4.14.4. Bin tane ağırlığı ... 244
4.14.5. Tanede protein ve sedimantasyon (M-SDS) oranı ... 248
5. SONUÇ VE ÖNERİLER ... 256
5.1. Sonuçlar ... 256
5.2. Öneriler ... 265
KAYNAKLAR ... 266
ix
SİMGELER VE KISALTMALAR
Simgeler
o
C gün Geçen termal gün sayısı r Korelasyon katsayısı
R2 Regresyon (determinasyon) katsayısı
Kısaltmalar
AÖF Asgari Önemli Fark (least significant difference, LSD 0,05 )
Bİ Başak İndeksi (tozlanma dönemi) BV Biyolojik verim, Biyokütle GDG Gelişme Derecesi-Gün Hİ Hasat İndeksi
KO Kareler Ortalaması KT Kareler Toplamı LAI Yaprak Alan İndeksi m2 Metrekare
mm Milimetre nm Nanometre Ö.D. Önemli Değil
SÇK Suda Çözünebilir Karbonhidrat SD Serbestlik Derecesi
SPAD Klorofil birimi VK Varyasyon Kaynağı
VK% Varyasyon Katsayısı (coefficient of variation, CV%) ve ark ve arkadaşları
1. GİRİŞ
Bitkisel üretimi sınırlandıran önemli faktörlerden biride kuraklıktır. Kuraklığın tanımlanan birçok çeşidi olmakla birlikte, dört belirgin kuraklık tipi olduğu kabul edilmektedir. Bunlar; meteorolojik kuraklık, tarımsal kuraklık, hidrolojik kuraklık ve sosyo-ekonomik kuraklıktır (Wilhite ve Glantz 1985). Yağışa bağlı iklim sınıflandırmalarında bazı kaynaklarca, yıllık ortalama yağışı 250 mm'den az olan yerlerin kurak, 250-500 mm arası olan yerlerin ise yarı kurak iklime sahip olarak tanımlandığı belirtilmekle birlikte, Türkiye’de kuraklık sınırı genellikle 400 mm olarak ifade edilmektedir (Kadıoğlu 2008). Brouwer ve Heibloem (1986)’da yaptıkları genel değerlendirmede, yıllık yağışı 100 mm’nin altında olan yerleri çöl, 100-400 mm arasında olanları kurak, 400-600 mm arasında olanları ise yarı kurak olarak sınıflamaktadır.
Buğdayın yetiştirildiği çoğu bölgelerde, özellikle Akdeniz ikliminde tane doldurma çeşitli fizyolojik ve biyolojik stres koşullarının etkisi altındadır. Bu dönemde çoğunlukla sıcaklık artar toprak nemi azalır, sapa kalkma döneminde stres koşulları karbon asimilasyonunu azaltır (Whan ve ark 1991). Saptaki kaynakların geliştirilmesi, çevresel ve biyotik stres faktörlerine karşı, tane doldurmayı destekleyen özelliği nedeniyle ıslahta önemlidir. Saplardaki deponun kapasitesinin büyüklüğü genetik yapısal özellikler tarafından kontrol edilmektedir ve saptaki birikim sapa kalkma ile birlikte gelişim karakteristiğine bağlı olarak bitkinin çok sayıda boğum oluşturması yüksek verim potansiyeli üzerine olumlu bir etki yapmaktadır (Blum 1998).
Tane doldurmada özümlemeye esas karbonun kaynağı, güneş ışınlarına maruz kalan yeşil yüzey alanıdır. Bu kaynak hem tane doldurma hem de bitkinin canlılığını devam ettirmesi için kullanılır. Çeşitli araştırıcılar tanenin yapısı ve gelişimi üzerine etkili olan karbonun üç kaynaktan sağlandığını belirtmektedir. Bunlar; fotosentezde biriktirilen, çiçeklenme öncesi ve çiçeklenme sonrası biriktirilen ürünlerin tekrar taşınması yolu ile olmaktadır. Çiçeklenme sonrası dönemde ışık alan yüzeyi fotosentez özümlemesi üzerine etkili iken kuraklık stresi bu dönemde fotosentezi azaltır, bu nedenle saptaki yedek depo ürünleri tane doldurma üzerine önemli bir kaynaktır (Kobata ve ark 1992, Schnyder 1993, Ehdaie ve Waines 1996, Blum 1998, Yang ve ark 2000). Çalışmalar göstermiştir ki saptaki Suda Çözünebilir Karbonhidrat’lar kuraklığa dayanımda fizyolojik özelliğin göstergesidir ve aynı zamanda stres koşullarında temel karbon kaynağı olarak tane doldurmaya katkı sağlar. Çünkü kuraklık gövdede bulunan
karbonhidratların taneye taşınmasını (translokasyon) ve etkinliğini artırır (Ehdaie ve ark 2006).
Kuraklığın beslenme üzerine olumsuz etkisi iyi bilinmektedir. Bununla birlikte kurak koşullarda yetiştirilen bitki çeşitleri arasında bir farklılık olup olmadığı ve bunun kuraklığa dayanıklılıkla bir ilgisinin olup olmadığı halen araştırılmaktadır. Besin etkinliği çok kompleks olup başlıca iki temel mekanizmayı içerir. Bunlar; besin maddesi alım etkinliği (kök salgıları ve kök morfolojisi tarafından kontrol edilen besin maddesi alım mekanizması) ve besin maddesi kullanım etkinliği (her bir gram besin maddesine karşılık üretilen kuru madde)'dir (El Bassam 1997).
Tane doldurma döneminde oluşan ani kuraklık ve yüksek sıcaklık streslerinde, çeşitlerin translokasyon kapasiteleri, verim kayıplarının azaltılması üzerine oldukça etkilidir. Çeşitlerin translokasyon kapasitelerinin ölçülmesi kurağa dayanıklılıkları hakkında bir gösterge olabilir. Ancak translokasyon kapasitesinin ölçülmesinde kullanılan kurutma (desikasyon) yöntemlerinin, uygulanması fazla zaman ve işçilik gerektirdiği için geniş materyal üzerinde çalışılmasını zorlaştırmaktadır.
Bu çalışmanın amacı; translokasyon oranını dolaylı yönden gösterecek sap doluluk oranı, saplardaki suda çözünebilir karbonhidrat miktarı ile translokasyon kapasitesi ve bunlar ile azot alım, kullanım etkinlikleri arasındaki ilişkileri araştırmaktır. Bu amaçla; Eskişehir koşullarında, 4 ekmeklik buğday çeşidi üzerinde, ilave azot gübrelemesinin yapıldığı, yağmura bağımlı ve takviye sulama koşullarında yürütülen denemelerle farklı sulama rejiminde tozlanma öncesi sapta biriktirilen ürünlerin tane doldurmaya etkisi araştırılmıştır. Bu çalışmayla, buğday verimini etkileyen temel unsurlar incelenirken, suyun kısıtlı olmadığı koşullar ve yağmura bağımlı koşullar birlikte ele alınıp, verim potansiyeli ve kurağa dayanıklılığı belirleyen bazı parametreler ile azot beslemesinin bu parametreler üzerine etkileri incelenmiştir. Çalışmada incelenen parametreler kısaca; biyolojik verim, rezerv translokasyonu, Suda Çözünebilir Karbonhidratlar (SÇK), azot alım etkinliği, sap doluluk oranı, bayrak yaprak klorofil okumaları (SPAD) ve yeşil alan sürdürme (stay green), başaklanma zamanı, bitki boyu, başak indeksi, hasat indeksi ve verim unsurlarıdır.
2. KAYNAK ARAŞTIRMASI
Turner (1982) translokasyonun başladığı dönemdeki biyolojik kütlenin rezerv kapasitesini belirleme yoluyla bu konuda etkili olduğunu bildirmiştir. Translokasyon kapasitesinin genel anlamda kurak koşullara uyumlulukla ilişkisini görmek amacıyla genotiplerin karşılaştırıldığı çalışmalarda çok farklı sonuçlar elde edilmesinden, bu özelliğin her türlü stres altında büyük yarar sağlamayabileceği, ancak genotiplerin sadece bu karakter açısından yapılan karşılaştırmalarda sonuç vermese bile (stepwise) çoklu regresyon türü yaklaşımlarla katkı payının araştırılabileceği belirtilmiştir. Kuraklıkla ilgili birçok özellikte olduğu gibi, bu konuda da parametreler arası interaksiyonların ilişkileri belirlemeyi zorlaştırdığı bildirilmektedir.
Blum ve ark (1983), buğdayda birçok karakterin çeşide özgü olarak farklılık gösterdiğini, bunlardan birinin de depolanmış karbonhidratın taneye taşınma (translokasyon) kapasitesi olduğunu bildirmişlerdir. Tane doldurma için gerekli olan, yaprak ve gövde de depolanmış karbonhidratın taneye taşınma hızı, süresi ve miktarı yönünden çeşitler arasında önemli farklılıklar bulunduğunu vurgulamışlardır.
Baker ve Gallagher (1983)’e göre, buğdayda kardeşler ana sapa ait yaprakların ekseninden çıkmakta, sap uzamasının başlamasından önce kardeşlenme bitmiş olmaktadır. Bununla birlikte diğer bir görüş ise; kardeşlenmenin belirli bir dönemde sona ermediği, kardeşlenmenin sona erdiği dönemin çeşide ait genetik özellikler ve çevre şartları tarafından belirlendiği yönündedir (Longnecker ve ark 1993).
Takahashi ve ark (1990) Hokkaido Üniversitesinde, üç yazlık buğday çeşidi ile yürüttükleri çalışmada, bitki sapını üst boğum (peduncle), ikinci (second internode) ve alt (basal) boğum olmak üzere üç kısımda, başaklanmadan olgunlaşma dönemine kadar 10 defa örnekleme yaparak suda çözünebilir karbondihrat (SÇK) miktarlarını incelemiştir. Çalışma sonucunda ikinci ve daha aşağıdaki boğumlarda SÇK miktarının birinci (peduncle) boğumdan daha yüksek olduğu ve bu kısımların ana depo organı oldukları, alt boğumlarda SÇK tozlanma döneminden önce birikirken birinci ve ikinci boğumda tozlanma dönemi ile başladığını bildirmişlerdir.
Davidson ve Chevalier (1990)’e göre çoğu kardeşler ana sapın uzaması sırasında ölürken, tane doldurma döneminde hemen hemen hiç kardeş ölümü gerçekleşmediğini belirtmektedir.
Van Oosterom ve Acevedo (1993) Suriye’de 1989 yılında iki farklı bölgede çevre ve fenolojinin arpanın gelişim ve verimi üzerine olan etkilerini araştırmak
amacıyla yaptıkları çalışmada; yaprakların yeşil kalma süreleri, özgül yaprak alanı ve bitki büyüme oranlarını erkenci, orta erkenci ve geçci çeşitlerde karşılaştırmışlardır. Çalışma sonucunda orta erkenci çeşitlerin erkenci çeşitlere göre daha uzun yaprakların yeşil kalma süresine sahip olurken en uzun yeşil kalmanın geçci çeşitlerde olduğunu ve bunun verimle zayıf korelasyon verdiğini, geçci çeşitlerin düşük bitki gelişme oranı ve düşük verim verdiğini, orta erkenci çeşitlerin geçci çeşitlere göre daha yüksek bitki gelişme oranı verdiğini ama erkenci çeşitlerden daha yüksek verim vermediğini, sonuç olarak yüksek bitki gelişme oranına ve yaprakların daha uzun yeşil kalma sürelerine sahip erkenci çeşitlerin bölge için uygun olacağını bildirmişlerdir.
Kalaycı ve ark (1998), Orta Anadolu’da kurağa dayanıklı buğday genotiplerinin belirlenmesi amacıyla 21 ekmeklik buğday çeşidi üzerinde bir çalışma yapmışlardır. Bu bölgede, en istikrarlı verime sahip çeşitlerin Gerek 79, Bolal 2973 ve Dağdaş olduğunu, normal düzeydeki kuraklıklarda özellikle yaprak eni ve bitki boyunun dayanıklılığı belirleyici en önemli morfolojik parametreler olduğunu, böylece uzun boylu ve dar yapraklı çeşitlerin bu düzeydeki kuraklıklara daha dayanıklı olduğunu, ancak sıcaklık stresi ile birlikte gelen aşırı kuraklık hallerinde fizyolojik parametrelerin önemli rol oynadığını belirtmişlerdir. Translokasyon kapasitesinin iri taneli çeşitler için daha önemli olduğunu vurgulamışlardır.
Blum (1998) tarafından fruktan sentezinde ilk adım ve belkide en önemlisinin fruktanın sakroz enzimi tarafından katalize edilmesi olduğunu belirtmiştir. SST (sucrose fructosyltransferase) aktivitesi substrat (sakroz) konsantrasyonu ile ilişkili olduğunu, buğdayda penultimate boğumda sakroz miktarı fazla olduğunda fruktan asimilasyonunun da arttığını, SÇK saptan harekete geçtiğinde (mobilize) toplam glikoz, sakroz ve fruktanın azaldığını fakat toplam fruktoz miktarı önce arttığı sonra azaldığını, bu durumun fruktanın kendi ürününden önce hidrolize olduğunu gösterdiğini belirtmiştir. Sapa kalkma döneminde stres koşulları karbon asimilasyonunu azaltmış ve yapılan çalışmada tekrar taşınabilir SÇK kuruda 641 mg, suluda 1047 mg bulunmuştur. Tarla koşullarında kuru alanlarda tane doldurma döneminde tekrar taşınabilir toplam SÇK miktarının, suludakine kıyasla ancak yarısının elverişli durumda olduğu, ortamdaki karbondioksit konsantrasyonu arttığında asimilasyonun arttığı ve saplarda daha fazla karbon biriktirildiği belirtilmiştir.
Blum (1998) geç dönemde oluşan kuraklık streslerinde verimi etkileyen en önemli faktörlerden birinin, bitkinin sap ve yapraklarında bulunan rezerv madde toplamı ile bitkinin bu maddeleri translokasyonla taneye taşıma kapasitesi olduğu, tanenin yapısı
ve gelişimi üzerine etkili olan karbonun üç kaynaktan sağlandığını belirtmektedir, bunlar; fotosentezde biriktirilen, çiçeklenme öncesi ve çiçeklenme sonrası biriktirilen ürünlerin tekrar taşınması yolu ile olduğu bildirilmektedir. Çiçeklenme sonrası dönemde ışık alan yüzey, fotosentez özümlemesi üzerine etkili bulunurken, kuraklık stresinin bu dönemde fotosentezi azalttığı bu nedenle saptaki yedek depo ürünlerinin tane doldurma üzerine etkili olduğu bildirilmiştir. Uygun koşullar altında tane doldurma döneminden önce sapta yeterli karbonhidrat birikiminin mümkün olduğu, çeşit özelliğine bağlı olarak tozlanma öncesi yeterli karbonhidrat birikiminin, yüksek verim potansiyelini desteklediği bildirilmiştir. Çalışmaları göstermiştir ki tane doldurmanın başlarında ve sararırken tekrar taşınmanın arttığı, saptaki SÇK’ların kuraklığa dayanımda fizyolojik özelliğin göstergesi olduğu ve aynı zamanda stres koşullarında temel karbon kaynağı olarak tane doldurmaya katkı sağladığı belirtilmiştir. Hasat indeksi ve kuru madde taşınımı arasında pozitif bir ilişki bulunduğu, uzun olgunlaşma dönemi ve yüksek kuru madde taşınımının su kullanım etkinliğini artırdığı ve bunların sonucu olarak hasat indeksi ve verimin arttığı bildirilmiştir.
Borrell ve ark (2000), Avustralya’da 8 adet hibrit sorgum çeşidi ile tane doldurma döneminde yaprakların yeşil kalma sürelerinin verime olan etkisini araştırmışlardır. Araştırıcılar sulu koşullarda yetişen bitkilerde yaprakların yeşil kalma sürelerinin verime herhangi bir etkisi olmazken, kurak koşullarda yaprakların uzun süre yeşil kalmasının toplam biyoması ve tane verimini olumlu yönde etkilediğini belirtmişlerdir.
Reynolds ve ark (2000), kuraklığa dayanıklı buğday geliştirilmesinde kullanılabilecek bir konsept model geliştirmişlerdir. Bu modelde yer alan parametrelerden; a) Tohumun iri olması; çıkış gücünü artırması, erken dönemde tarla yüzeyini kapatmaya yardımcı olması nedeniyle başlangıç biyolojik veriminin yüksek olmasına yardım ederken. b) Uzun koleoptil; derin ekimlerde çıkış gücünü artırdığı, köklerin derindeki suya ulaşmasına yardım ederek ve toprak yüzeyinde oluşan yüksek sıcaklığın çimlenmeyi etkilemesini önlediğini bildirmiştir. c) Erken dönemde tarla yüzeyini kapatması; ince ve geniş yapraklı çeşitler tarla yüzeyini daha iyi kapattığı ve böylece toprak suyunun muhafazası ve radyasyon kullanım etkinliğinin yükselmesini sağladığı bildirilmektedir. Bu özellik bilhassa yağışların erken dönemde geldiği Akdeniz iklim kuşağında kurağa dayanıklılık açısından daha önemli bulunmuştur. d) Tane doldurma öncesi yüksek biyolojik verim; tane doldurma öncesi yüksek biyolojik verim sahip çeşitlerin su kullanım etkinliğinin daha yüksek olduğu bildirilmiştir. e)
Translokasyon kapasitesinin yüksek olması; tane doldurma döneminde oluşan stres koşullarında saplardan taneye taşınan depo besin maddeleri önemli olduğundan bu kapasitenin yüksek olmasını sağlayacak olan kalın ve uzun sapların depo besin maddeleri açısından gerekli olduğu vurgulanmıştır. f) Başak fotosentez kapasitesinin yüksek olması; başakların su kullanım etkinliğinin yapraklarınkinden daha yüksek olduğu ve yapmış olduğu fotosentez ürünlerinin taneye katkısı stres koşullarında %40’a kadar çıktığı bildirilmektedir. g) Kılçıklılık; başak fotosentezi açısından önemli katkı sağladığı, h) Yapraklarda yüksek oransal nem içeriği, stoma iletkenliği ve düşük bitki örtüsü sıcaklığı; bu fizyolojik parametrelerin bitkinin topraktan daha iyi su aldığını gösteren endirekt göstergeler olduğu bildirilmektedir. ı) Ozmotik düzenleme; yaprakların düşük su potansiyeline sahip olması kurak koşullarda bitkinin su alımına yardım ettiği, i) ABA birikimi; kurak koşullarda biriken ABA miktarı bitkinin stoma iletkenliği, hücre bölünmesi, bitki organlarının büyüklüğüne bağlı olduğunu, ancak fazla miktarda ABA birikimi bitkide çiçek sterilitesine neden olabildiği bildirilmektedir. j) Yüksek sıcaklığa dayanıklılık, yaprak anatomisi; mumsuluk, tüylülük, yaprak kıvrılması, kalınlık; bitkinin aşırı radyasyon yüklenmesini engellediği ve evapotranspirasyon oranının düşmesine neden olarak kuraklıktan dolayı geri dönüşümsüz zararlanmaları engellediği bildirilmektedir. Ancak bu özelliklerin aşırı olması optimum koşullarda radyasyon kullanım etkinliğini azaltması nedeniyle verimi kısıtlayabildiği vurgulanmakta, k) Kardeşlerin hayatiyetini devam ettirebilme özelliğinin yüksekliği, y) Yaprakların yeşil kalma süresinin uzunluğu gibi yukarıda açıklanan parametrelerin kurağa dayanıklı buğday geliştirilmesinde seleksiyon kriteri olarak kullanılabileceğini bildirmişlerdir.
Cseuz ve ark (2002), Macaristan’da buğday ıslah programında tarla koşullarında kuraklığa dayanıklı hatların seleksiyonunda kullanılabilecek olan translokasyon kapasitesi ve bayrak yaprağı oransal nem içeriklerini karşılaştırdıkları çalışmada; bayrak yaprağına tane doldurmanın 14. günü % 2’lik NaClO3 uygulamışlar ve bu yapraklarda
karbonhidrat ölçümlerinin yanı sıra oransal nem içeriklerini belirlemişlerdir. Oransal nem içeriğinin daha çok kütikula dayanıklılığı ile alakalı olduğunu, bu nedenle diğer parametrelerle ilişki vermediğini, translokasyon kapasitesinin ise seleksiyon kriteri olarak kullanılabileceğini ama yine de morfolojik parametrelerin seleksiyonda hala önemli olduğunu bildirmişlerdir.
Yang ve ark (2003), iki ekmeklik buğday çeşidiyle yaptıkları çalışmada, iki farklı N dozu uygulaması yanı sıra tam sulama ve başaklanmadan 9 gün sonra kuraklığa
tabi tutulan bitkilerde; bayrak yaprağında fotosentez etkinliği, klorofil, ABA, Zeatin ve Zeatin riboside ölçümü yapmışlardır. Çalışma sonucunda; bayrak yaprağı fotosentez oranı ve klorofil miktarının kuraklık uygulanan bitkilerde hızlı bir düşüş gösterdiğini, bunun sonucunda kuraklık stresinin yapraklarda sararmayı artırdığını, kuraklığın gövdede bulunan karbonhidratların taneye taşınmasını artırdığını, tane doldurma süresini kısalttığını, bunun yanında tane doldurma oranını artırdığını, yapraklarda ve gövdede ABA konsantrasyonunu artırırken Zeatin ve Zeatin riboside konsantrasyonunu düşürdüğünü, ABA konsantrasyonunun bayrak yaprağı fotosentez oranı ve klorofil içeriği ile istatistiki olarak negatif korelasyon verirken Zeatin ve Zeatin riboside ile pozitif korelasyon verdiğini, yüksek ABA miktarının translokasyonu ve tane doldurma oranını artırdığını, yapraklara kinetin uygulanmasının ise tam tersi etki yaptığını, ABA ve sitokininin yapraklardaki sararmayı kontrol mekanizmasının içinde yer aldıklarını bildirmişlerdir.
Berry ve ark (2003), metrekarede sap sayısının en yüksek 1000–1600 olduğu ve bu oluşan kardeşlerin %32 - %63’ünün öldüğünü belirtmektedir. Kardeş ölümlerinin çeşitlere göre değişmekle birlikte en yoğun yaşandığı dönemlerin, bayrak yaprak çıkış dönemi (ZD 39) ve çiçeklenme dönemi (ZD 61) olduğunu belirtmişlerdir. Kardeşlenme potansiyeli açısından genotipler arasında büyük farklılıklar olduğu gibi çevre koşulları ve kültürel uygulamalar da maksimum kardeş sayısını etkilediği, birçok konuda olduğu gibi, maksimum kardeş sayısının belirlenmesinde de en etkili çevre unsurları su ve sıcaklık olduğu, kardeşlenmenin su sıkıntısına karşı hassas olduğu ve çok kurak koşullarda yarıya kadar düşebileceği belirtilmektedir (Rickman ve ark 1983, Peterson ve ark 1984).
Verma ve ark (2004), fotoperiyoda hassas Beaver buğday çeşidi ile hassas olmayan Soissons buğday çeşidinin melezlemesinden oluşan doubled haploid popülasyonda bayrak yaprak sararmasının kuraklıkla ve verimle alakasını araştırdıkları çalışmada; farklı dönemlerde bayrak yaprağındaki yeşil alan miktarını ölçmüşler ve aynı zamanda bu özelliği kontrol eden genleri belirlemek için genetik haritalama yapmışlardır. Yapmış oldukları değerlendirmede; farklı çevre koşullarında bayrak yaprağı yeşil kalma süresi ile verim arasında pozitif bir korelasyon olduğunu, sararma sırasında 2B ve 2D kromozumu arasında bir çatışma olduğunu bu nedenle çok kompleks bir yapıya sahip olduğunu bildirmişlerdir.
Gong ve ark (2005), tane doldurma döneminde fotosentez oranı ve translokasyonla taşınan karbonhidrat miktarlarını hibrit ve normal ekmeklik buğday
çeşitlerinde karşılaştırdıkları çalışmada; bitki beslenmesini takip etmek için işaretli
14
CO2 kullanmışlardır. Araştırma sonucunda hibrit çeşitte 14CO2’ün bayrak yaprağına
taşınmasının normal çeşitten daha yavaş olduğunu, tane doldurma dönemindeki bayrak yaprağındaki fotosentez oranının ve klorofil içeriğinin hibrit çeşitte normal çeşide oranla çok daha yavaş düştüğünü, bunun sonucunda tane doldurma süresinin normal çeşide oranla 6 gün daha uzun sürdüğünü, hibrit çeşidin daha düşük hasat indeksine sahip olduğunu, sap kısmında kalan depo karbonhidrat miktarının hibrit çeşitte daha fazla olduğunu, hibrit çeşitte verim potansiyelini artırmak için hasat indeksinin yükseltilmesinin temel amaç olması gerektiğini bildirmişlerdir.
Ehdaie ve ark (2006), sulu ve kurak koşullarda iki yıl süre ile yürüttükleri çalışmada 11 buğday çeşidinde boğumlar arası uzunluk, ağırlık ve çeşitlerin sap özgül ağırlığının karbonhidrat birikimi ve translokasyona olan etkilerini araştırmışlardır. Çeşitler arasındaki farklılığı belirlemek amacıyla 10 gün arayla karbonhidrat birikimi ve translokasyon oranlarını belirlemişlerdir. Çalışma sonucunda; boğum arası mesafelerin, ağırlığının ve özgül ağırlığının kurak koşullarda azaldığını, üst boğumdan taşınan depo besin maddelerinin sulu koşullarda kuru koşullardakine göre daha az olduğunu, buna karşın orta ve alt boğumdan taşınan maddelerin daha fazla olduğunu, kuraklığın translokasyon etkinliğini artırdığını, taşınan toplam maddenin yüzdesi olarak ifade edildiğinde bunun üst boğumda %65, orta boğumda %11 ve alt boğumda ise %5 olduğunu, sapın maksimum özgül ağırlığının translokasyona uğrayan madde miktarı ile alakalı olduğunu, buğdayda bitki boyunda üst boğum, orta boğum ve alt boğumda dengeli bir uzunluk dağılımının depo besin maddelerinin gövdede birikimi ve translokasyon açısından önemli olduğunu bildirmişlerdir.
Ehdaie ve ark (2006) diğer çalışmalarında, kuraklığın orta ve alt boğumda SÇK içeriğini ve konsantrasyonunu azalttığını, sulu koşullarda orta ve alt boğumdan taşınan SÇK’nın kurak koşullara oranla daha fazla olduğu, kurak koşulların SÇK translokasyon etkinliğini artırdığını ve bu artışın en fazla üst boğum aralığında olduğunu, tane doldurma sırasında ölçülen SÇK içeriğinin farklı boğum aralıklarından taşınan toplam SÇK ile yüksek korelasyon verdiğini, denemede yer alan makarnalık buğdayın bütün boğum aralıklarında en yüksek SÇK içeriğine sahip olduğunu ve sonuç olarak boğumların SÇK içeriğinin ölçülmesinin kuraklığa dayanıklı buğday geliştirmede seleksiyon kriteri olarak kullanılabileceğini belirtmişlerdir.
Zhang ve ark (2006), tarla koşullarında yetiştirilen iki yazlık ekmeklik buğday çeşidinin bayrak yapraklarında fotosentetik özellikleri inceledikleri çalışmada; bayrak
yaprağın çıkışından 10 gün sonra her iki çeşitte de maksimum fotosentez oranının gözlendiğini ve bu noktanın sararmanın başlangıcı olduğunu, çıkıştan 27 gün sonra fotosentezde hızlı bir düşüş olduğunu, NM9 çeşidinin NM8 çeşidine oranla daha fazla klorofil içerdiğini ve daha yüksek klorofil a/b oranına sahip olduğunu, klorofildeki azalmanın diğer çeşide oranla daha yavaş olduğunu ve bunun sonucunda bayrak yaprağının yeşil kalma süresinin daha uzun sürdüğünü ve bunun daha yüksek verim vermesinin nedeni olduğunu bildirmişlerdir.
Çekiç (2007), Eskişehir’de 30 buğday çeşidiyle yaptıkları çalışmada, tane doldurma döneminde oluşan kuraklık stresi bayrak yaprakların daha erken dönemde sararmasına neden olduğunu bildirmiştir. Yıllar arasında ortalama bayrak yaprak yeşil kalma süreleri karşılaştırıldığında ise yağışların daha fazla olduğu yılda daha uzun olduğu, 2 yılın ortalaması olarak, kuru koşullarda %35.7 olan translokasyon oranı, sulu koşullarda % 18,0 olarak gerçekleştiği bildirilmiştir.
Çekiç (2007), bayrak yaprak yeşil kalma süresinin kurak koşullara uyumlulukla ilişkisi üzerine Eskişehir’ de yaptığı bir çalışmada, Fischer ve Maurer (1978)’e göre hesaplanan kurak hassasiyet indeksleriyle en yüksek ilişkiyi veren parametrenin bayrak yaprak yeşil kalma süreleri olduğunu bildirmiştir. Bu özellikde, tane doldurma süresi gibi, başaklanma tarihiyle çok yakın ilişkili bulunmuş ve kuraklığa terkedilen parsellerde R2 = 0,96** gibi yüksek düzeyde ilişki verirken, sulanan parsellerde ilişkinin önemsiz çıkması sonucunda bu özelliğin kuraklık koşullarında önemli bir parametre olarak değerlendirilebileceği belirtilmiştir.
Önder (2007) tarafından 2005–2006 yetişme periyodunda, kuru (yağmura bağımlı) ve destek sulamalı şartlarda, 10 farklı ekmeklik buğday çeşidi ile (Ak-702, Kıraç-66, Bezostaja 1, Gerek-79, Kutluk-94, Süzen-97, Aytın-98, Harmankaya-99, Altay-2000, Sönmez-2001) Eskişehir’de yürütülen çalışmasında; beş farklı gelişme döneminde (Zadoks ve ark 1974), kardeş sayıları, biyokütle ağırlıkları, başak indeksi (tozlanma dönemi), hasat indeksi ve verim öğeleri incelenmiştir. Çalışmada yer alan çeşitlerin kuru şartlarda; maksimum kardeşe sahip olduğu dönemde (Zadoks 23) bitki başına kardeş sayıları 2.06 ile 3.36 arasında değişirken, kardeş ölüm oranları %50 ile %73 arasında değiştiği belirtilmiştir. Bayrak yaprak dönemine (Zadoks 41) kadar kardeş ölümleri yönünden sulu ve kuru arasında bir fark bulunmazken esas farklılık bu dönemden sonra başladığı ve sulamanın fertil kardeş sayısını artırdığı belirtilmiştir. Verim üzerine etkili olan diğer morfolojik ve fizyolojik parametreler incelendiğinde; kuru koşullarda sapa kalkmaya kadar olan biyokütle ölçümleri, verimle ilişki
bulunurken, çok kardeşlenen çeşitten ziyade kardeşlerin hayatiyetini özellikle kuru koşullarda en iyi muhafaza eden çeşitlere gidilmesi uygun görüldüğü belirtilmektedir.
Farshadfar ve ark (2008), kuraklığa dayanımın moleküler indekslerini değerlendirmek amacıyla 20 farklı ekmeklik buğday çeşidi üzerinde sulu ve kuru koşullar altında yürüttükleri tarla çalışmalarında; bayrak yapraklarda analizler yapılmış, tane verimi ile prolin, SÇK ile toplam protein (yaprakta) arasında pozitif korelasyon bulmuşlardır. SÇK ile; verim 0.82**, protein 0.57**, prolin 0.75** seviyelerinde pozitif ilişkiler bulunmuş ve bu parametreleri yüksek olan genotiplerin kurağa mukavemet gösterebilen çeşitler olduğu belirtilmiştir.
Ercoli ve ark (2008), buğday yetiştirildiği dönem itibarıyla tozlanma dönemine kadar yeterli yağışı alabildiği, tane doldurma dönemi başladığında kısıtlı yağmur ve yükselen sıcaklıkların etkisi ile su stresi oluştuğunu bildirmiştir. Tane doldurma süresince tanede biriktirilen karbonhidratlar iki kaynaktan geldiği birincisi ürünlerin (asimilant) doğrudan taneye gönderilmesi, ikincisi yedek depolarda biriktirilen ürünlerin tekrar taşınması şeklinde olduğunu bildirmiştir. Çalışmada; iki makarnalık buğday çeşidi ile 12 kg N/da ve 18 kg N/da azot seviyelerinde, dört farklı sulama rejiminde (tam suluma, düşük stres, orta stres ve yüksek stres), iki yıl süren çalışma sonucunda; tozlanma döneminde azot ve çeşitler test edilmiş çeşitler arasında farklılık görülmezken azot seviyeleri farklılık oluşturmuştur. İlave azot tüm bitki organlarında, kuru ağırlıkta (g/m2), azot içeriğinde (g/kg) ve azot kapsamında (kg/da) önemli artışlar sağlamıştır. Birim alandaki başak sayısı sulamadan etkilenmemiş, azot uygulamalarından kısmen etkilenmiş, başakta tane sayısı ve tane ağırlığı sulama ve azottan etkilenmiş, artan azot olumsuz etkiyi azaltmıştır. Artan su stresi koşullarında azot verilmeyen parseller hariç azot verilen parsellerde kök kuru ağırlığı artmıştır. Hasat olgunluğuna geldiğinde artan azot ve artan su stresi ile birlikte tüm bitki organlarında (yaprak, sap, kök ve kavuz) azot içeriği artmıştır. Yine hasat döneminde, artan su stresi ile birlikte tane azot kapsamı azalmış ancak yaprak, sap ve kökte azot kapsamı artmıştır. Tane doldurma süresince kuru madde birikimi azottan olumlu etkilenirken, su stresinden olumsuz etkilenmiştir. Artan azot ile birlikte, azotun tekrar taneye taşınma oranı ve etkinliğinin arttığı bildirilmiştir.
Moayedi ve ark (2009), 4 makarnalık ve 1 ekmeklik buğday çeşidinde dört farklı sulama rejiminde tozlanma öncesi sapta biriktirilen ürünlerin tane doldurmaya etkisini araştırdıkları çalışmada, 5-9-5 NPK/da ve 7 kg N/da ilkbahar gübresi olarak verilmiş, kuru madde translokasyonu (KMT) sulama rejimlerinden etkilendiği vurgulanmıştır.
Bununla birlikte kuru madde translokasyon etkinliği (KMTE) ve tozlanma öncesi asimilantların taneye katkısı (TÖAK) yönünden genotip ve genotip çevre interaksiyonu önemli olduğu belirtilmiştir. En yüksek KMT 836 mg/bitki ile ekmeklik buğdayda bulunurken, en düşük makarnalık buğdayda 521 mg/bitki bulunmuştur. Kurak koşullara gidildikçe TÖAK oranı artarken, KMTE’de “genotip x çevre” interaksiyonu anlamlı olmuş, çeşitler kuru madde translokasyon etkinlikleri yönünden kuraklığa karşı farklı tepkiler vermiştir. Bazı çeşitlerin TÖAK oranı kuraklık arttıkça arttığı ve bu çeşitlerin ileride kuraklığa dayanıklı genotip olarak kullanılabileceği düşünülmüştür. Çünkü saptaki SÇK’lar fotosentezin sınırlandığı stres koşullarında tane verimini destekleyen karbon kaynağı olduğu, başaklanma süresi uzun ve uzun boylu genotiplerin ideal nem koşullarında diğer genotiplere göre başaklanma öncesi sahip oldukları yedek karbonhidratlarını artırdığı, başak kuru ağırlığı ve başak hasat indeksinin tane verimine esas teşkil eden depolar olup bu deponun büyüklüğünün verimi olumlu etkilediği bildirilmiştir.
Farklı araştırıcılar tarafından, fotosentez ve tane oluşumu için optimal sıcaklığın 20 oC dolayında olduğu, bunun üzerindeki sıcaklıkların fotosentezi olumsuz etkilediği (Camp ve ark 1982, Al-Khatib ve Paulsen 1984) ve bunun taneye asimilat aktarımını azalttığı (Wardlaw ve ark 1980) bildirilmiştir. Çiçeklenme dönemindeki 30 oC ve üzeri sıcaklıklarda ise tamamen sterilite oluştuğu bildirilmiştir (Owen 1971, Saini ve Aspinall 1982). Bu şekilde yüksek sıcaklık, kuraklık veya başka stres unsurlarının etkisiyle yapraklar kurumaya başlarken, bunun da translokasyonu tetiklediği ve depolanmış karbonhidratların taneye taşınmaya başlandığı açıklanmıştır (Blum 1998).
Taşınmayı başlatan sinyalin kaynağı tam olarak bilinmemekle birlikte, gibberellinler ya da absisik asit (ABA) gibi hormonların bu konuyla ilgili olabilecekleri tahmin edilmektedir (Yang ve ark 2001). Stres olmayan koşullarda da translokasyonun önemli katkı yapabileceği açıklanmış (Gebbing ve Schnyder 1999) ancak en büyük katkısının stres koşullarında olduğu bildirilmiştir (Blum 2005). İdeal koşullarda, tane doldurmada kullanılan karbonhidratların %95’inin o dönemdeki karbon assimilasyonuyla sağlandığı belirtilmiş (Kobata ve ark 1992), ancak stres oluştuğunda, translokasyonla aktarılan rezerv oranlarının arttığı bildirilmiştir.
3. MATERYAL VE YÖNTEM
3.1. Materyal
Bu çalışmada, Geçit Kuşağı Tarımsal Araştırma Enstitüsü Müdürlüğü tarafından tescil ettirilen kışlık ekmeklik buğday çeşitleri Müfitbey (M), Bezostaja 1 (B), İzgi 2001 (İ) ve ES 26 (E) materyal olarak kullanılmıştır.
Eskişehir koşullarında, iki yıl, ilk yıl tek yerde, sulu (S), kuru (K) olmak üzere iki set halinde yürütülmüştür (Şekil 3.1). İkinci yıl denemeler iki yerde yürütülmüş olup; iki set sulu (S1 ve S2), iki set kuru (K1 ve K2) olmak üzere (Şekil 3.2), toplam üç yerde yürütülen çalışmada varyasyon kaynağı olarak bir kontrol ve üç farklı azot dozu kullanılmıştır. Bu azot dozları sırasıyla; 0 kg N/da (Kontrol), 5 kg N/da, 10 kg N/da ve 15 kg N/da’dır. Azot dozlarının yarısı ekimle birlikte, kalan yarısı kardeşlenme döneminde Amonyum Nitrat (% 33 N) şeklinde verilmiştir. Toprak analiz sonuçlarına göre, ekimle birlikte 7 kg P2O5/da fosfor TSP ile her parsele uygulanmıştır.
Şekil 3.1. Birinci yıl (2010-2011) genel tarla planı
3.2. Deneme Deseni
Tesadüf Bloklarında Bölünmüş Parseller deneme deseninde, ana parseller; azot dozları, alt parseller; çeşitler olmak üzere, 4 tekerrürlü ve 2 set (destek sulu ve yağmura bağımlı koşullarda) olarak yürütülmüştür.
Bir set; 4 (azot) x 4 (çeşit) x 4 (tekerrür) = 64 parselden oluşmaktadır. Buna göre bir set yağmura bağımlı (kuru), bir set destek sulu olmak üzere iki set toplam 64 x 2 = 128 parselden oluşmaktadır. Parsel boyutları ekimde; 7 m x 1.2 m (6 sıra, 0.2 m), hasatta; 5 m x 1.2 m alınmıştır. Her iki set için uygulanan deneme planı Çizelge 3.1’de verilmiştir.
Çizelge 3.1. Bir set (sulu/kuru) için deneme deseni ve faktörlerin dağılımı
Azot Dozu N15 N 15 N 15 N 15 N 5 N 5 N 5 N 5 N 10 N 10 N 10 N 10 N 0 N 0 N 0 N 0 Çeşit İ E M B İ E M B İ E M B İ E M B 4.Tekerrür 4 Azot Dozu N 0 N 0 N 0 N 0 N 10 N 10 N 10 N 10 N 15 N 15 N 15 N 15 N 5 N 5 N 5 N 5 Çeşit B M E İ B M E İ B M E İ B M E İ 3.Tekerrür 3 Azot Dozu N 5 N 5 N 5 N 5 N 15 N 15 N 15 N 15 N 0 N 0 N 0 N 0 N 10 N 10 N 10 N 10 Çeşit E İ B M E İ B M E İ B M E İ B M 2.Tekerrür 2 Azot Dozu N 10 N 10 N 10 N 10 N 0 N 0 N 0 N 0 N 5 N 5 N 5 N 5 N 15 N 15 N 15 N 15 Çeşit M B İ E M B İ E M B İ E M B İ E 1.Tekerrür 1
*Müfitbey (M), Bezostaja 1 (B), İzgi 2001 (İ), ES 26 (E)
3.3. Deneme Yerinin Meteorolojik Özellikleri
Gelişme Derecesi-Gün (GDG) değerini hesaplayabilmek için günlük en düşük ve en yüksek sıcaklıklar ile aylık yağış miktarları kaydedilmiştir.
GDG = [( Tmak + Tmin)/2] – Tbaz (3.1)
Burada;
Tmak ve Tmin günlük maksimum ve minimum sıcaklıkları (oC),
Tbaz buğdayın fizyolojik faaliyetlerinin devam ettiği minimum sıcaklığıdır. Bu çalışmada Tbaz 4.4 oC
olarak alınmıştır.
Çalışmanın yürütüldüğü yer olan Eskişehir’in iklimi incelendiğinde uzun yıllar yağış ortalamasının 400 mm’nin altında 348.5 mm olarak gerçekleşmiştir (Çizelge 3.2).
Çalışmanın yürütüldüğü yıllarda yağış toplamı sırasıyla 443.6 mm ve 374.6 mm olarak gerçekleşmiştir.
Çizelge 3.2. Enstitü arazisinde yapılan yağış ölçümleri (mm)
Yıllar E yl ü l E ki m K as ım A ra lı k O ca k Ş uba t M art N is an M ayı s H az ir an T em m u z A ğus tos Toplam (mm) Uzun yıl* 14.6 26.2 30.0 45.7 38.5 32.4 33.4 35.2 43.7 28.7 13.7 6.4 348.5 2005-06 15.0 13.4 41.5 20.7 53.5 29.1 25.2 8.9 23.5 12.0 37.5 0.0 280.3 2006-07 58.4 46.8 13.3 7.8 45.5 16.5 39.7 19.4 36.9 20.4 20.0 0.0 324.7 2007-08 0.0 19.2 92.4 49.9 15.7 1.0 42.4 38.5 11.7 9.3 0.0 5.5 285.6 2008-09 30.7 6.4 49.6 34.5 66.3 82.0 40.9 28.0 15.4 10.2 19.4 2.0 385.4 2009-10 7.1 9.0 29.5 65.1 36.0 42.8 32.6 23.9 20.7 79.0 7.4 0.9 354.0 2010-11 22.5 77.1 7.5 60.4 26.6 8.9 20.0 56.9 145.8 9.4 8.5 0.0 443.6 2011-12 2.1 57.9 0.0 46.1 58.0 42.1 56.4 22.1 80.9 0.0 5.5 3.5 374.6 *Uzun yıl G.K.T.A.Enstitü’sünde 1926-2012 yılları arasında yapılan ölçümlerden hesaplanmıştır.
Geç dönem kuraklık stresi üzerinde etkili olan Mart, Nisan ve Mayıs aylarından oluşan ilkbahar yağışları incelendiğinde; uzun yıllarda 112.3 mm iken denemenin birinci yılında 222.7, ikinci yılında 159.4 mm olarak gerçekleşmiştir.
Birinci yıl ilkbaharda görülen yüksek yağış destek amaçlı yapılan sulamanın (S) etkinliğini azaltmış ve hatta yüksek azot dozlarında çeşitlere göre farklı oranlarda yatmaya neden olmuştur. Ayrıca yağmura bağımlı, kuru koşullarda (K) oluşması beklenen ilkbahar geç dönem kuraklık stresinin yeterince oluşmasını engellemekle birlikte, bu riski ortadan kaldırmak ve tarla koşullarında suni olarak kuraklık oluşturmak amacıyla parsellere desikant uygulaması yapılmış ve translokasyon oranları bu uygulamalar üzerinden hesaplanmıştır.
Yağışla birlikte kuraklık üzerine etkili olan diğer bir faktör olan hava sıcaklığı günlük olarak izlenmiş (Çizelge 3.3) ve ekim tarihinden sonra 4.4 oC (Tb) üzerinde geçen günlerin sıcaklık toplamları alınarak Gelişme Derecesi Gün (GDG) değerleri hesaplanmıştır.
Çizelge 3.3. Farklı gelişme dönemlerinde yapılan örnekleme tarihleri ve GDG değerleri Gelişme dönemi (Zadoks’a göre) Örnekleme tarihi (Birinci yıl) GDG ZD 31 02 Mayıs 2011 413 ZD 41 17 Mayıs 2011 504 ZD 71 09 Haziran 2011 765 ZD 94 25 Temmuz 2011 1429 Gelişme dönemi (Zadoks’a göre) Örnekleme tarihi (İkinci yıl) GDG ZD 31 04 Mayıs 2012 300 ZD 41 17 Mayıs 2012 437 ZD 71 06 Haziran 2012 643 ZD 94 13 Temmuz 2012 1235
Çizelge 3.3 incelendiğinde bitkiler Zadoks 31 (birinci boğumun görüldüğü) dönemine her iki yılda da mayıs ayının ilk haftası ulaşırken, gelişme derecesi gün (GDG) olarak incelendiğinde bu değer birinci yıl 413 ikinci yıl 300 birim olarak gerçekleşmiştir. Bu durumda araştırmanın ikinci yılında, ekimden birinci boğum dönemine kadar geçen kış ve ilkbahar dönemlerinin diğer yıla göre daha serin geçtiğini göstermektedir.
3.4. Tarla Denemeleri Ekim ve Hasat Zamanları
Tarla denemeleri, Geçit Kuşağı Tarımsal Araştırma Enstitüsü arazisinde; Tesadüf Bloklarında Bölünmüş Parseller deneme deseninde, Ana Parseller; azot dozları, Alt Parseller; çeşitler olmak üzere, 4 tekerrürlü ve 2 set (destek sulu ve yağmura bağımlı koşullarda) iki yıl yürütülmüştür.
Birinci yıl; bir yağmura bağımlı (K) ve bir destek sulama (S) koşullarında iki set halinde, 25.10.2010 tarihinde ekilmiş ve 25.07.2011 tarihinde hasat yapılmıştır (Şekil 3.1).
İkinci yıl; iki yağmura bağımlı (K1 ve K2) ve iki destek sulama (S1 ve S2) koşullarında (Şekil 3.2) dört set halinde, 01.11.2011 tarihinde ekilmiş ve 13.07.2012 tarihinde hasat yapılmıştır.
Şekil 3.3. 22 Kasım 2010 tarihinde denemelerden genel görünüm
Şekil 3.4. Kardeşlenme döneminde azotun ikinci yarı uygulaması ve genel görünüm
Denemede yer alan 0, 5, 10 ve 15 kg N/da azot dozlarının yarısı ekimle birlikte verilmiş olup, kalan yarısı kardeşlenme döneminde Amonyum Nitrat (% 33 N) şeklinde her parsel için ayrı ayrı tartılarak uygulanmıştır. Ekimle birlikte 7 kg P2O5/da fosfor
TSP gübresi ile uygulanmıştır.
3.5. İstatistiki Analiz
Tesadüf Bloklarında Bölünmüş Parseller deneme deseninde, ana parseller; azot dozları, alt parseller; çeşitler olmak üzere, 4 tekerrürlü ve 2 set (destek sulu ve yağmura bağımlı koşullarda) olarak yürütülen bu çalışmada elde edilen veriler, Çizelge 3.4’te görülen model kullanıllarak, setler ayrı analiz edilmiştir. Ortalamaların gruplandırılmasında Student’s t yöntemiyle (Student 1908) elde olunan 0.05 önemlilik düzeyindeki AÖF değerleri kullanılmıştır. Korelasyon (r) hesaplamaları Pearson (1920) (Pearson product-moment correlation)’a göre yapılmıştır.
Çizelge 3.4. Verilerin değerlendirilmesinde kullanılan varyans analiz modeli
Varyasyon Kaynağı SD SD
Tekerrür t-1 3
Azot dozu a-1 3
Hata (txa) txa 9
Çeşit ç-1 3
Azot dozu x Çeşit axç 9
Hata (a-1) x (ç-1) x t 36
Toplam n-1 63
3.6. Denemelerde Alınan Ölçüm ve Gözlemler
3.6.1. Translokasyon kapasitesi
Kimyasal uygulaması tozlanmadan yaklaşık olarak 2 hafta sonra yapılmakta ve potasyum iyodür (KI) veya magnezyum klorat (Mg(ClO3)2) gibi oksitleyiciler
kullanılmak suretiyle klorofil molekülleri parçalanarak yeşil aksam kısa sürede kurutulmaktadır. Aynı parsellerin işlem yapılmamış kısmındaki başaklarda oluşan taneler kontrol olarak alınıp, uygulama yapılan bitkilerde translokasyonla oluşan tane ağırlığı kontrolle karşılaştırılmak suretiyle translokasyon oranları hesaplanmıştır.
Başaklanmadan 20-25 gün sonra parsellerin bir kısmına bitki yeşil kısımlarını kurutmak amacıyla desikant (%4’lük Magnezyum Klorat (Mg(ClO3)2) tüm bitki
aksamı ıslatılıncaya kadar uygulanmıştır. Bitkiler bu uygulamadan 2-3 gün sonra kurumuş, geri kalan kısmı kontrol olarak bırakılmıştır.
Hasat zamanına kadar beklenerek bu dönemde kontrol ve uygulama yapılan bölümden başak örneği toplanmıştır.
Toplanan başak örnekleri 80 ºC’de 24 saat kurutulduktan sonra taneler başaktan ayrılarak tartılmıştır.
Translokasyon kapasitesi Blum ve ark (1983) ve Kalaycı ve ark (1998) tarafından bildirilen aşağıdaki eşitliğe göre hesaplanmıştır;
TK% = (USD-UÖD)/(KD-UÖD)x100 (3.2)
Burada: TK=Translokasyon kapasitesi, USD =Uygulama sonrası bir tane ağırlığı (mg), UÖD = Uygulama öncesi bir tane ağırlığı (mg), KD=Kontrol uygulamasında bir tane ağırlığı (mg)
3.6.2. Sapta, suda çözünebilir karbonhidratlar
* (Yemm ve Willis 1954)
Şekil 3.6. SÇK’ların belirlenmesinde kullanılan yönteme* ait süreç
Öğütülüp tartılan sap örneklerinden 0.2 g örnek alınıp 50 ml saf su eklenerek, 20
o
C’de 1 saat çalkalanmıştır. Çalkalandıktan sonra santrifüj edilerek sıvı ve katı kısım birbirinden ayrılır. Reaksiyon için 200 µl örnek alınıp, üzerine 10 ml anthrone reagent* eklenmiştir. Kaynayan (100 oC) su banyosunda 10 dakika bekletilip, sıcak su banyosundan alınan örnekler buzlu su ile soğutulduktan sonra Spektrofotometrede 620 nm dalga boyunda okunmuştur (Şekil 3.6). Hazırlanan standart glikoz çözeltileri Şekil 3.7’de görülmektedir.
*Anthrone reagent Hazırlanışı; 500 ml konsantre H2SO4, 200 ml deiyonize su ile seyreltilir, bu
seyreltilmiş çözeltiden 100 ml alınır ve 0.2 g anthrone eklenir.
0.2 g Anthrone 100 ml H2SO4 500 ml H2SO4 + 200 ml H2O ANTHRONE REAGENT 0.2 g Örnek 50 ml H2O 20 0C 1 saat çalkala Santrifüj . 200 µl 10 ml Ant.Rea. Çalkala 100 0 C 10 dk Su banyosu Buzlu su ile soğut 620 nm Spektrofotometre oku
Şekil 3.7. Spektrofotometrede okunmak üzere hazırlanmış 0-35 ppm standart glikoz çözeltileri
3.6.3. Kuraklık hassasiyet indeksi
Kuraklık hassasiyet indeksi (KHİ) Fischer ve Maurer (1978)’a göre aşağıdaki şekilde hesaplanmıştır.
Kuraklık şiddeti = (Sulu ort. verim*- Kuru ort. verim*)/Sulu ort. verim (3.3) KHI = [(Sulu verim** – Kuru verim**)/Sulu verim**]/Kuraklık şiddeti (3.4)
*Burada kullanılan veriler denemede yer alan tüm çeşitlerin ortalamasıdır. **Kullanılan veriler bir çeşidin verim değerleridir.
3.6.4. Bayrak yaprak yeşil kalma süresi
Farklı tarihlerde SPAD metre (Şekil 3.8) ile okunan SPAD değerleri bağımlı değişken (y), okumanın yapıldığı tarihlerdeki başaklanma tarihinden itibaren Gelişme Derecesi Gün (GDG) değerleri ise bağımsız değişken (x) olarak alınmak sureti ile kuatratik regresyon yöntemi ile klorofilin sıfıra düştüğü teorik nokta hesaplanarak bayrak yaprak yeşil kalma süresi GDG cinsinden hesaplanmıştır (Çekiç 2007, Lopes ve Reynolds 2012).
3.6.5. Yaprak alan indeksi
Yaprak Alan İndeksi (LAI) ölçümlerinde, “LAI-2000” marka optik sensör (Şekil 3.9) kullanılmıştır. Sensör yapısı gereği okuma anında LAI değerlerini daha sonra kullanılmak üzere depolamakta, hesaplanmış indeks değerleri olarak sunmaktadır. Cihaz kendi ışık kaynağına sahip olmadığı için atmosferden gelen ışık miktarı doğrudan etkili olmaktadır, bu nedenle okumalar, ışık yoğunluğunun fazla değişiklik göstermediği sabahın erken saatlerinde yapılmıştır. 74 derecelik görüş açısına sahip sensörde, okumalar esnasında 90 derecelik perde kullanılmıştır. Sensörün çalışma prensibi gereği bir kez bitki örtüsü üzerinden okuma (A) yapılırken, bitki örtüsü altından daha fazla sayıda okuma (B) yapılır, sensör görüntüsü halka şeklinde eş merkezli beş açıya bölünmüştür (Welles ve Cohen 1996) ve her açı için bir değer okunmaktadır, bu halkalar bitki örtüsü üzerinde tutulan veya yansıtılan radyasyon ile bitki örtüsü altına geçen radyasyon (490 nm altında) arasındaki orana göre her açıya ait B değerlerini, kendine ait A değerine bölünmesi ile hesaplanır (Welles ve Norman 1991). Her parselde bir üst (A) okuması ve bitki örtüsü altından dört adet alt (B) okuma yapılmıştır.
Şekil 3.9. LI-COR, LAI-2000 Plant Canopy Analyzer, optik el sensörü
3.6.6. Biyokütle ağırlığı
Biyokütle içindeki mevcut rezervin taneye taşınması, tane doldurmaya katkıda bulunup, kuraklık stresinden oluşan zararı hafifletici olabilmektedir.
Reynolds ve ark (2005)’a göre; kardeş sayılarındaki farklılıklar bitki örtüsünü ve ışıktan yararlanma oranlarını da etkiler, kardeş sayılarının hesaplanabilmesi için en az üç farklı dönemde sayım yapılmalıdır (Şekil 3.10); kardeş sayısının maksimum olduğu dönem (sapa kalkma dönemi başlangıcı, bu dönemde tüm kardeşler sayılır), tozlanma dönemi (tüm canlı kardeşler sayılır) ve olgunluk dönemi (sadece başaklı kardeşler sayılır).
Şekil 3.10. Buğdayın gelişimi ve büyüme dönemlerinin şematik diyagramı
E=ekim; Çi=çimlenme; Ç= çıkış; TYK= toprak yüzeyini kaplama; BDB= başaklanma dönemi başlangıcı; B= başaklanma; T= tozlanma; TDDB= tane doldurma dönemi başlangıcı; FO= fizyolojik olgunluk; BD= büyüme dönemi. Slafer ve Rawson (1994)’dan alınmıştır.
Reynolds ve ark (2001)’a göre; biyokütle örnekleri 4 farklı bitki gelişim döneminde (Zadoks ve ark 1974)’a göre alınmıştır, bu dönemler sırası ile; ZD 31 (sapa kalkma dönemi), ZD 41 (Bayrak yaprak dönemi), ZD 71 (Süt olum dönemi), ZD 94 (hasat olgunluğu) dönemleridir.
Biyokütle örnekleri için sıra üzerinde 50 cm’lik bir alan biçilerek naylon poşet içerisine konulduktan sonra, nem kaybını önlemek amacıyla, poşet ağzı sıkıca kapatılıp, zaman kaybedilmeden laboratuarda tartılıp, 75 oC’de 48 saat bekletilmek üzere etüve konulmuş ve kuru ağırlıkları tartılmıştır.
Alınan bu biyokütle örneklerinden; kardeş sayıları, biyolojik verim (biyokütle), bin tane ağırlığı, metrekarede tane sayısı, metrekarede başak sayısı, başakta tane sayısı, hasat indeksi ve başak indeksi değerleri hesaplanmıştır.
Biyokütle örneklemesi için alınan örneklerde, Reynolds ve ark (2001)’a göre; Toplam yaş ağırlık (TYA) tartıldıktan sonra,
Alt örnekler seçilir, Alt örnek yaş ağırlığı (AÖYA) ve 75 oC’de 48 saat bekletilerek, Alt örnek kuru ağırlığı (AÖKA) tartılmıştır.
Başaklanma döneminden sonra yapılan örneklemelerde başak indeksi (Bİ) değerini hesaplayabilmek için bitki örnekleri alınır, Başaklar (B), Sap ve Yapraklarından (SY) ayrılarak her ikisinin fırın kuru ağırlıkları tartılmıştır.
Bu işlemler sonucunda;
Biyokütle (g/m²) = TYA x (AÖKA/AÖYA)/alan (3.5)
m² sap (kardeş) sayısı = Biyokütle/KA (1 kardeşin kuru ağırlığı) (3.6)
Bİ = BKA/SYKA (3.7)
Verim öğelerinin hesaplanması için ise bitkiler hasat olgunluğuna geldiğinde;
Parseli temsil edecek bir alandan kesilip alınan bitkiler içerisinden 50 adet başaklı bitki (BB 50) seçilip, kese kağıtları içine konularak 75 0C ‘de 48 saat kurutulmuştur (KA 50).
Bu seçilen 50 bitki hasat indeksinin (Hİ) hesaplanabilmesi için harman edilerek, tane ve saplar ayrılmıştır.
Parsel veriminin bulunması için tüm parsel hasat edilmiştir. 250 tane sayılarak ağırlıkları tespit edilmiştir.
Bu işlemler sonucunda;
Hİ = Tane ağırlığı (BB 50)/Toplam kuru ağırlık (KA 50) (3.8)
Biyokütle (g/m²) = Verim/Hİ (3.9)
BTA (g) = 250 tane ağırlığı x 4 (3.10)
Tane sayısı (m²) = Verim/BTA x 1000 (3.11)
Başak sayısı (m²) = Biyokütle/Sap kuru ağırlığı (tek bitki) (3.12) Tane sayısı/Başak = Tane sayısı (m²)/Başak sayısı (m²) (3.13)
3.6.7. Tozlanma öncesi asimilantların tekrar taşınması (remobilization)
Arduini ve ark (2006)’a göre:
KMT (mg/bitki) = KMS t - KMS h (3.14)
KMTE % = [KMT/KMS t] x 100 (3.15)
TÖAK % = (KMT / TV) x 100 (3.16)
KMT; (Kuru Madde Translokasyonu), KMS t; (Kuru Madde Sapta, tozlanma dönemi), KMS h; (Kuru Madde Sapta, hasat dönemi), KMTE; (Kuru Madde Translokasyon Etkinliği), TÖAK; (Tozlanma Öncesi Asimilatların Taneye Katkısı), TV (tane verimi mg/bitki)
Robertson ve Giunta (1994)’a göre;
Başak Hasat İndeksi % = BKA (tozlanma döneminde)/SKA (tek bitki hasatta) (3.17)
ATT = Tüm bitki azot kapsamı (tozlanmada)–saplar azot kapsamı (hasatta, tane hariç) (3.18)
ATTE % = (ATT/Tüm bitki azot kapsamı (tozlanmada)) x 100 (3.19)
ATTK % = (ATT/Tane azot kapsamı (hasatta)) x 100 (3.20)
AHİ % = (Tane azot kapsamı/Bitki azot kapsamı ) x 100 (3.21)
ATT (Azot Tekrar Taşıma), ATTE (Azot Tekrar Taşıma Etkinliği), ATTK (tanede azot kapsamı üzerine Azot Tekrar Taşınmasının Katkısı), AHİ (Azot Hasat İndeksi), BKA (başak kuru ağırlığı), SKA (Sap kuru ağırlığı)
3.6.8. Bitki boyu
Her parselde, bitkinin toprak yüzeyinden son başakcığının üst noktasına kadar olan mesafe cm olarak ölçülmüştür. Kılçık yükseklikleri dahil edilmemiştir (Pask ve ark 2012).
3.6.9. Üst boğum (peduncle) uzunluğu
Bitki boyu ölçümü sırasında üst boğum ile başağın ilk başakcık boğumu arasındaki uzunluk alınırak, her tekerrürde parsellerin ortalarındaki 5 adet bitkinin ana sapında en üst boğum arası ölçülmüştür. Ölçüm yapılan 5 bitkinin aritmetik ortalaması alınarak her parsel için üst boğum arası uzunluğu (cm) tespit edilmiştir (Pask ve ark 2012).
3.6.10. Sap doluluk oranı
Tozlanma döneminde 5 bitkide orta boğumun (penultimate), yatay kesit alanı kumpas ile ölçülerek (Şekil 3.11) aşağıdaki gibi hesaplanmıştır.
Dolu kısım alanı (cm2) = Dış çap alanı (cm2) – İç çap alanı (cm2) (3.22) Hacim (cm3) = Dolu kısım alanı (cm2) x bitki boyu (cm) (3.23)
3.6.11. Başaklanma-olum süresi
Başaklanma tarihleri her parseldeki toplam başakların %50’den fazlasının, başak kınından %50 oranında çıktığı zaman dikkate alınarak tespit edilmiştir. 1 Mayıs’tan itibaren takvim günü olarak belirlenip, çeşitlerin tane doldurma süresince gösterecekleri klorofil değişimlerini de takip edebilmek amacıyla GDG olarak hesaplanmıştır.
3.6.12. Tane verimi
Başlangıçta 7 m uzunluğunda ekimi yapılan parsellerin 5 m’lik kısmı, tane verimi için “Hege” marka özel parsel biçerdöveri ile hasat edilmiştir. Her parsel (1.2 m x 5.0 m) 6.0 m² olarak hasat edilip, elde edilen parsel verimi, kg/da birimine çevrilmiştir.
4. ARAŞTIRMA SONUÇLARI VE TARTIŞMA
4.1. Translokasyon Kapasitesi
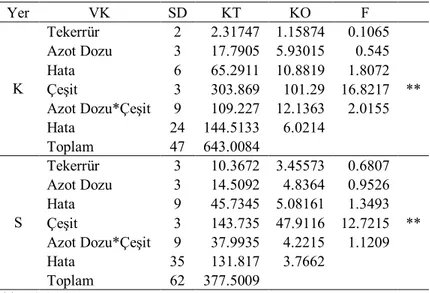
Araştırmanın birinci yılında belirlenen translokasyon oranına ait varyans analiz sonuçları Çizelge 4.1’de, bu özelliğe ait ortalama değerler ise Çizelge 4.2’de verilmiştir.
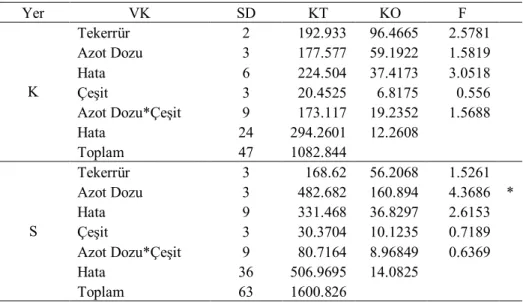
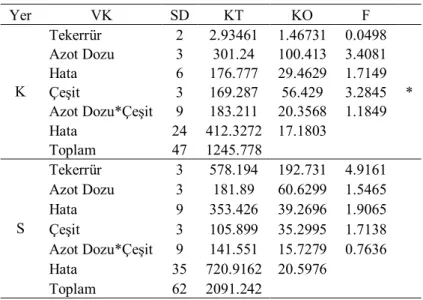
Çizelge 4.1. Birinci yıl kuru (K) ve sulu (S) koşullarda translokasyon oranlarına ait varyans analizi
Yer VK SD KT KO F K Tekerrür 2 211.681 105.84 0.7189 Azot Dozu 3 1981.56 660.52 4.4778 * Hata 6 882.628 147.105 0.945 Çeşit 3 4094.7 1364.9 8.7679 * Azot Dozu*Çeşit 9 5648.64 627.627 4.0318 * Hata 18 2802.051 155.67 Toplam 41 16215.15 S Tekerrür 3 948.704 316.235 2.1682 Azot Dozu 3 179.094 59.6981 0.3954 Hata 9 1280.79 142.31 0.568 Çeşit 3 4576.28 1525.43 6.0888 * Azot Dozu*Çeşit 9 4000.85 444.539 1.7744 Hata 26 6513.795 250.531 Toplam 53 17360.35 * P<0.05
Çizelge 4.2. Birinci yıl kuru (K) ve sulu (S) koşullarda translokasyon oranlarına ait ortalamalar
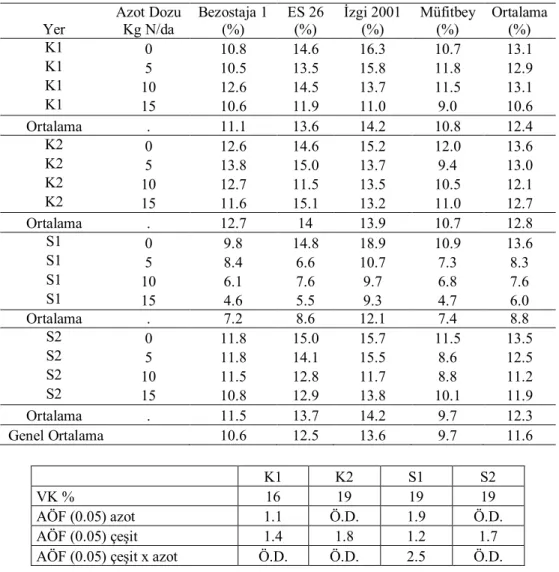
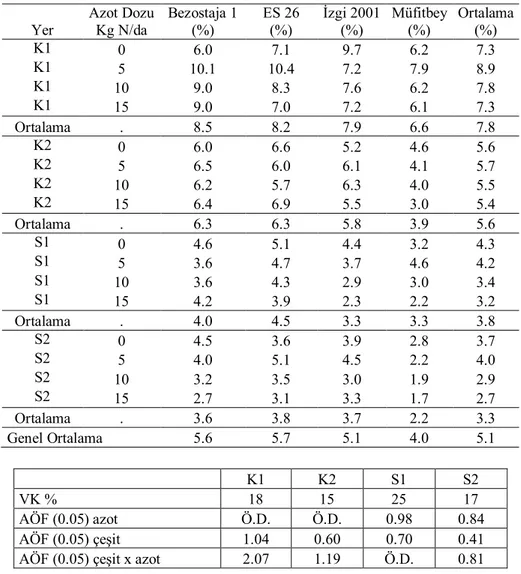
Yer Azot Dozu Kg N/da Bezostaja 1 (%) ES 26 (%) İzgi 2001 (%) Müfitbey (%) Ortalama (%) K 0 19 12 21 34 22 K 5 21 18 21 31 23 K 10 26 62 32 44 41 K 15 35 22 5 72 34 Ortalama . 25 29 20 45 30 S 0 17 15 23 55 28 S 5 31 26 15 29 25 S 10 32 12 15 30 22 S 15 43 33 1 42 30 Ortalama . 31 22 14 39 27 Genel Ortalama . 28 25 17 42 28 K S VK % 40 56 AÖF (0.05) azot 14.1 Ö.D. AÖF (0.05) çeşit 10.7 13.7
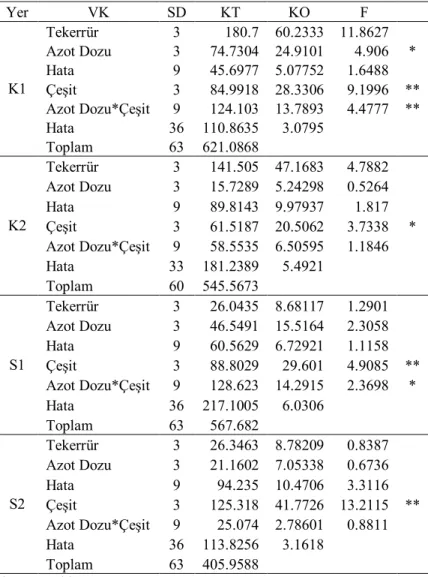
Elde edilen verilere göre İlkbahar geç dönem kuraklık stresinin yaşandığı yıllarda, tane doldurmaya destek veren ve kök, sap ve yapraklarda depolanan fotosentez ürünlerinin taneye taşınması (translokasyon) çeşitler ve azot dozları arasında farklı oranlarda gerçekleşmiştir. Çizelge 4.1 incelendiğinde birinci yıl, kuru koşullarda azot uygulamaları translokasyon oranları üzerine etkili olurken sulu koşullarda etkili olmamıştır. Çeşitler arasındaki farklılık her iki koşulda da önemli olmuştur. İlk yıl Müfitbey çeşidi %42 translokasyon oranı ile ilk sırada yer alırken İzgi 2001 %17 ile son sırada yer almıştır. Çeşitler ortalaması olarak, translokasyon oranı kuru koşullarda 10 kg N/da azot düzeyinde %41’e, sulu koşullarda 15 kg N/da azot düzeyinde %30’a çıkmıştır (Çizelge 4.2). Araştırmanın ikinci yılında belirlenen translokasyon oranına ait varyans analiz sonuçları Çizelge 4.3’de, bu özelliğe ait ortalama değerler ise Çizelge 4.4’de verilmiştir.
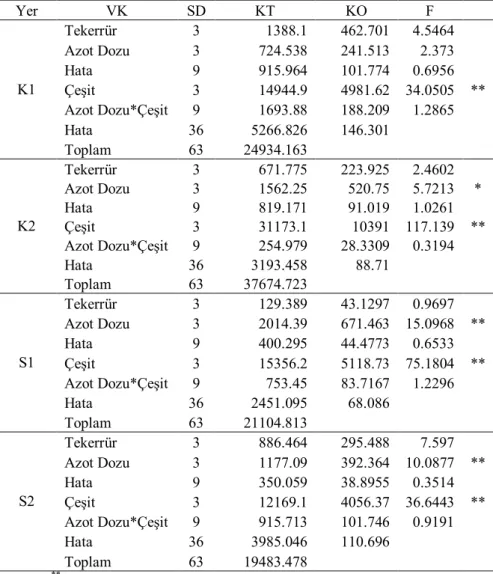
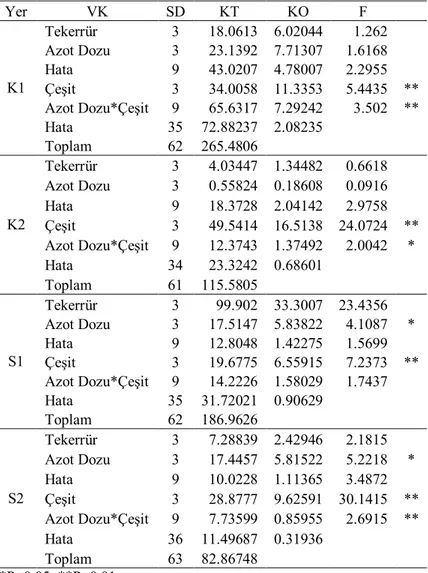
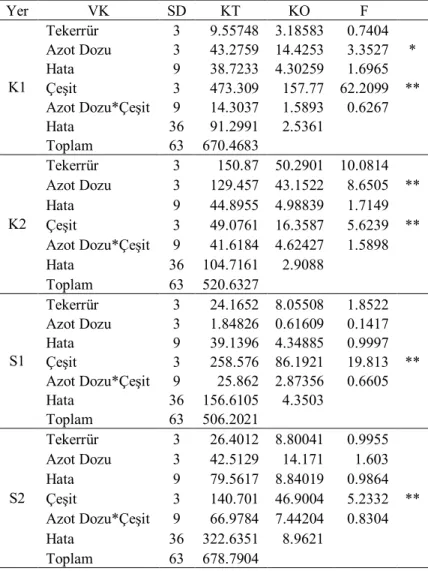
Çizelge 4.3. İkinci yıl kuru (K) ve sulu (S) koşullarda translokasyon oranlarına ait varyans analizi
Yer VK SD KT KO F K1 Tekerrür 3 1388.1 462.701 4.5464 Azot Dozu 3 724.538 241.513 2.373 Hata 9 915.964 101.774 0.6956 Çeşit 3 14944.9 4981.62 34.0505 ** Azot Dozu*Çeşit 9 1693.88 188.209 1.2865 Hata 36 5266.826 146.301 Toplam 63 24934.163 K2 Tekerrür 3 671.775 223.925 2.4602 Azot Dozu 3 1562.25 520.75 5.7213 * Hata 9 819.171 91.019 1.0261 Çeşit 3 31173.1 10391 117.139 ** Azot Dozu*Çeşit 9 254.979 28.3309 0.3194 Hata 36 3193.458 88.71 Toplam 63 37674.723 S1 Tekerrür 3 129.389 43.1297 0.9697 Azot Dozu 3 2014.39 671.463 15.0968 ** Hata 9 400.295 44.4773 0.6533 Çeşit 3 15356.2 5118.73 75.1804 ** Azot Dozu*Çeşit 9 753.45 83.7167 1.2296 Hata 36 2451.095 68.086 Toplam 63 21104.813 S2 Tekerrür 3 886.464 295.488 7.597 Azot Dozu 3 1177.09 392.364 10.0877 ** Hata 9 350.059 38.8955 0.3514 Çeşit 3 12169.1 4056.37 36.6443 ** Azot Dozu*Çeşit 9 915.713 101.746 0.9191 Hata 36 3985.046 110.696 Toplam 63 19483.478 * P<0.05; **P<0.01
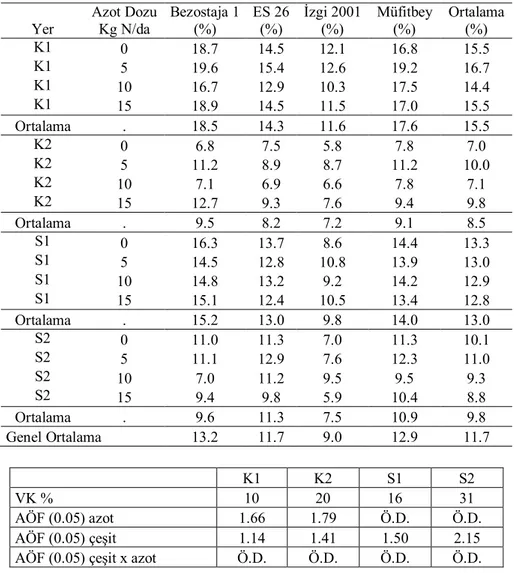
Çizelge 4.4. İkinci yıl kuru (K) ve sulu (S) koşullarda translokasyon oranlarına ait ortalamalar Yer Azot Dozu Kg N/da Bezostaja 1 (%) ES 26 (%) İzgi 2001 (%) Müfitbey (%) Ortalama (%) K1 0 27.2 5.9 9.2 43.3 21.4 K1 5 30.5 7.5 7.0 46.3 22.8 K1 10 42.0 11.5 7.0 51.8 28.1 K1 15 47.5 13.1 21.7 35.1 29.4 Ortalama . 36.8 9.5 11.2 44.1 25.4 K2 0 36.8 5.3 8.8 51.1 25.5 K2 5 40.0 6.9 9.2 60.4 29.1 K2 10 43.8 8.8 10.3 64.8 31.9 K2 15 50.3 18.0 15.9 71.8 39.0 Ortalama . 42.7 9.8 11.1 62.0 31.4 S1 0 29.9 6.2 7.6 41.5 21.3 S1 5 31.4 11.5 9.5 51.4 26.0 S1 10 32.6 13.3 12.1 42.0 25.0 S1 15 35.1 26.1 18.3 66.2 36.4 Ortalama . 32.2 14.3 11.9 50.3 27.2 S2 0 29.4 24.1 11.4 37.6 25.6 S2 5 31.9 20.4 12.5 51.0 29.0 S2 10 37.6 24.9 16.9 52.6 33.0 S2 15 36.8 26.8 18.0 66.6 37.1 Ortalama . 33.9 24.0 14.7 51.9 31.1 Genel Ortalama 36.4 14.4 12.2 52.1 28.8 K1 K2 S1 S2 VK % 48 30 30 34 AÖF (0,05) azot Ö.D. 7.6 5.3 5.0 AÖF (0,05) çeşit 8.7 6.7 5.9 7.5
AÖF (0,05) çeşit x azot Ö.D. Ö.D. Ö.D. Ö.D.
İkinci yıl (Çizelge 4.4) translokasyon oranı %52.1 ile en yüksek Müfitbey çeşidinde gerçekleşirken, en düşük %14.7 ile İzgi 2001 çeşidinde olmuştur. İkinci yılda hem kuruda hem suluda artan azot uygulamalarının translokasyon oranları üzerine olumlu etki yaptığı görülmektedir.
İzgi 2001 ve ES 26 başaklanma tarihi itibariyle erkenci çeşitler olup translokayon oranları orta geççi çeşitler olan Bezostaja 1 ve Müfitbey’den daha düşüktür. Bu durum tane doldurmaya katkısı yönünden geççi çeşitlerde translokasyonla taşınan fotoasimilant miktarının erkencilere göre daha önemli olduğunu ve ayrıca erkenci çeşitlerde fotosentez süresinin daha uzun olması nedeniyle translokasyonla taşınan miktarın daha az etkili olduğunu göstermektedir (Şekil 4.1). Benzer şekilde bazı araştırmacılar; erkenciliğin daha önemli olduğu (Blum ve Pnuel 1990), yüksek sıcaklığın translokasyonu artırıcı özelliğine neden olarak, sıcaklık baskısı altında, nişasta oluşumunda etkili nişasta sentez enziminin aktivitesini önemli ölçüde yitirmesi