http://journals.tubitak.gov.tr/agriculture/ © TÜBİTAK
doi:10.3906/tar-1112-2
Changes in mycorrhizal spore and root colonization of coastal dune vegetation of the
Seyhan Delta in the postcultivation phase
Özlem AYTOK1, K. Tulühan YILMAZ1, İbrahim ORTAŞ2,*, Halil ÇAKAN3
1 Department of Landscape Architecture, Faculty of Agriculture, University of Çukurova, 01330 Adana, Turkey 2 Department of Soil Science and Plant Nutrition, Faculty of Agriculture, University of Çukurova, 01330 Adana, Turkey
3 Department of Biology, Faculty of Science and Art, University of Çukurova, 01330 Adana, Turkey
1. Introduction
The Mediterranean dunes, which range from mobile to fixed dunes, have been overexploited. Indigenous dune forests were cleared and were temporarily cultivated or overgrazed by sheep or goats (van der Meulen and Salman 1996). Similarly, the coastal dune ecosystem of the Seyhan Delta, about 3200 ha of which was cultivated, is under risk of detrimental impacts of agricultural practices. Yılmaz (2002) investigated the impacts of agriculture, afforestation, and recreation on the sand dunes of the Çukurova deltas, including the Seyhan and Ceyhan deltas. Flora and some vegetation characteristics of the Çukurova deltas have been already examined by Çakan et al. (2005) and Yılmaz (2002). In comparison to the detailed studies on the flora and vegetation, the relation between plant communities and mycorrhizal richness and its presence in the plant communities on the sand dunes has received relatively little attention in the Mediterranean part of Turkey. Therefore, Çakan and Karataş (2006) investigated
the mycorrhizal status of dune plant species in relation to their life forms along a successional gradient of natural sand dunes.
Coastal sand dunes are ideal for investigating the roles of mycorrhiza in succession, as it is possible to examine vegetation through all serial stages, from the pioneer to the stabilized zones, in a relatively small area (Çakan and Karataş 2006). Mycorrhizal colonization has great importance on plant establishment and growth in sand dune ecosystems (Sylvia 1989; Sylvia and Jarstfer 1997, Ramos-Zapata et al. 2011). Previous research on the mycorrhiza of sand dunes has detailed the mycorrhizal status of the colonizing plants and the identification, distribution, and abundance of arbuscular mycorrhizal (AM) spores (Corkidi and Rincon 1997; Ramos-Zapata et al. 2011). It has also been demonstrated that the external mycelium of AM plays a significant role in the process of dune stabilization, as sand grains are bound together mechanically by their hyphae (Koske 1975; Sutton Abstract: Changes in the abundance of arbuscular mycorrhizal (AM) spores and root colonization that occurred in the postcultivation
phase were investigated in the coastal dunes of the Seyhan Delta, Turkey. A sampling method was used to obtain community-level information, which is essential for the evaluation of relations between plant communities and AM colonization. Eight quadrats of 10 × 10 m were selected to describe floristic composition of different plant communities. TWINSPAN was applied to identify the plant communities. In addition, DECORANA was employed to put forward a clear ordination of the communities. Soil samples were collected at depths of 0–20 cm from each quadrat, and site conditions were described by means of soil analyses such as extractable P (NaHCO3), pH, organic matters, and salinity (%). Differences were detected in the floristic composition of the vascular plant cover in quadrats located in the abandoned peanut-cultivated fields when compared with control quadrats. This difference occurred with an increase of weeds and decrease of native psammophytes in terms of species richness and relative coverage. Higher AM colonization rates were found in the abandoned fields than in the undisturbed sand dunes. These rates were associated with the amount of P2O5 and organic matters. It was concluded that the increase in those rates was a result of overfertilization and introduction of weeds. Transport of manure into dune fields enhances the potential for the introduction of weed seeds and results in further development of mycorrhizal symbiosis through rapid weed colonization. AM colonization rate (%) was detected to be much higher on fibrous and rhizomatous roots than tap and bulbous roots, respectively. The highest values of AM colonization were detected on the species belong to the family Gramineae.
Key words: Dune vegetation, synanthropic flora, mycorrhiza, agriculture
Received: 01.12.2011 Accepted: 23.05.2012 Published Online: 15.01.2013 Printed: 15.02.2013 Research Article
and Sheppard 1976; Clough and Sutton 1978; Forster 1979; Forster and Nicolson 1981a, 1981b). Mycorrhizal colonization also benefits soil rehabilitation and erosion control by stimulating soil aggregation (Abbott et al. 1992; Ortaş 2002).
Pioneer plants in coastal sand dunes are short of nutrients (N, P, K) and organic matters, and they are subject to wide fluctuations in soil moisture and temperature, injury from sand blasting and salt spray, and excessive erosion or accretion (Barbour et al. 1985; Maun 1994; Ramos-Zapata et al. 2011). Although it has been suggested that mycorrhizae are of great ecological significance to plants in sand dunes to increase nutrient uptake (Koske and Polson 1984; Read 1989) where the scarcity of phosphorus, nitrogen, and potassium can be extreme, this statement has not been experimentally verified and few studies have reported the influence of AM on the growth of sand dune species (Koske and Gemma 1995; Little and Maun 1996; Beena et al. 2000; van den Koornhuyse et al. 2003). Previous studies showed a negative correlation between the intensity of land use and species richness of sand dune vegetation (Acosta et al. 2000). The species composition, dominant life forms, and AM colonization rates in the sand dune vegetation also vary with time (Çakan and Karataş 2006). Ramos-Zapata et al. (2011) reported that the density of spores was higher during the rainy compared to the dry season. The present study deals with changes in mycorrhizal spore and root colonization of coastal dune vegetation following peanut cultivation on the dunes of the Seyhan Delta.
2. Materials and methods 2.1. Study site
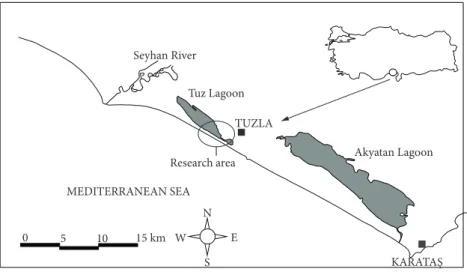
The study site is located near the Mediterranean coast of Adana Province, Turkey. Fieldwork was carried out on the
left bank of the Seyhan River estuary, in the town Tuzla in the Karataş district (36°40′N–36°44′N, 34°59′E–35°06′E) (Figure 1). The climate is typical semiarid Mediterranean, with a dry summer and a rainy winter. The average annual precipitation is 677 mm and the average temperature is 18.63 °C. The Seyhan River basin shows diverse landscape features and covers an area of 20,600 km2. Along the Seyhan Delta, extensive sand dunes and 2 lagoons, namely Tuz Lagoon and Akyatan Lagoon, form a mosaic structure of habitats at the coastal edge of the province (Yılmaz et al. 2004).
Sand dunes (mobile or fixed), sandy beaches, salt flats, salt marshes, riversides, fresh water marshes, field margins, and roadsides were classified as the principal habitats of the area (Çakan et al. 2005). Related to the habitat diversity, the floristic composition of the coastal area is very diverse. According to Çakan et al. (2005), the total native flora of the Çukurova deltas consists of 600 plant taxa. Sixty-two threatened plant taxa in the total flora (including 31 endemic and 31 rare taxa for Turkey) were designated as having the most restricted distribution and as being under threat of extinction.
2.2. Vegetation analysis
Quadrat sampling was performed to understand the relations between species composition of different communities and AM colonization. Eight quadrats (10 × 10 m) approximating the mean minimum area with prevailing plant communities were selected in foredune, humid dune slack, and salt pioneer sward locations. Of those 8 quadrats, 3 were in abandoned fields, 4 were in undisturbed dunes as control quadrats, and 1 represented a seminatural site. The plant species within each quadrat were recorded and collected for identification, following the nomenclature of Davis (1965–1988). During field works, the species richness (i.e. the number of species)
Tuz Lagoon Akyatan Lagoon MEDITERRANEAN SEA Research area TUZLA N 5 W E S 15 km KARATAŞ Seyhan River 0 10
and relative coverage were recorded. Root samples for each species present in each quadrat were collected to detect mycorrhizal status from both the undisturbed sand dune sites and the abandoned fields.
To explain causal relations between AM colonization and agriculture, the classification used by Yılmaz (2002) was followed. It enables one to characterize natural dune vegetation (NDV) and synanthropic vegetation (SV) in habitats created or influenced by man and livestock.
2.3. Root sampling, mycorrhizal colonization, and soil analysis
Root samples were obtained from 52 plant species collected from the quadrats in foredunes, humid dune slacks, and salt pioneer swards locations. Roots were separated from the core soil following the method of Ortas et al. (1996). These samples were preserved in a mixture of ethanol, acetic acid, and formalin and were cut into 1-cm-long pieces to determine the root length colonization by AM. Therefore, 50 root segments from 3 individuals of each species occurring in each quadrat were obtained. The root cleaning and staining procedure were performed following the method described by Koske and Gemma (1989). The percentage of AM root colonization was calculated as the number of 1-cm-long infected root segments out of the 50 contained on slides (Giovannetti and Mosse 1980). The number of spores in a 5-g sand sample inoculate mixture was counted using the wet sieve extraction method (Gerdemann and Nicolson 1963). Soil samples were collected at depths of 0–20 cm from each quadrat and were stored in polyethylene bags for transport to the laboratory. The edaphic properties of the soils, such as organic matter, salinity, pH, and phosphorus, were determined according to the methods of McLean (1982) and Murphy and Riley (1962).
2.4. Statistical analyses
Two-way indicator species analysis (TWINSPAN) (Hill 1974) was used to identify the plant communities; similarly, detrended correspondence analysis (DCA) was employed to obtain an effective ordination of plant communities and their relation along the natural and land use gradients. To understand the correlation between site conditions, such as organic matter, salinity, pH, phosphorus in soil, and vegetation characteristics such as life forms and plant communities, and the data on AM colonization, findings were analyzed using the Statistical Analysis System (SAS 1987).
3. Results 3.1. Vegetation
During the floristic investigation, 52 plant species were identified. They constituted 24 perennial, 22 annual, and 6 biennial species. The total coverage of perennials was recorded as 15% in 8 quadrats, whereas the coverage of
annuals and biennials was determined as 35%. Relative coverage of the annuals was determined to be 50% in the seminatural quadrat and 48% where agricultural practices were prevalent. The rate of total NDV species in undisturbed sites (quadrats 2, 3, 6, and 7) was 83%. The total coverage and species number of NDV was higher than SV in the quadrats where the sand dunes were undisturbed. The highest coverage of NDV (53%) was observed at quadrat 3, where species diversity was very poor. In quadrat 1, vegetation cover was higher than the total average and species richness had the maximum value (25 species).
Occurrence of each species and mean AM root colonization rates of the TWINSPAN groups are shown in Figure 2. These groups were compared with syntaxonomic classifications, covering units of coastal vegetation. Group A and group B were well matched with Euphorbio-Ammophiletea arundinaceae J.M. & J. Géhu 1988, whereas group C was matched with Arthrocnemetea fruticosi Tx. & Oberd. 1958, which was the representative of the halophytic communities occurring in the study site.
The most common species in the study area was Cyperus capitatus Vandelli. Parapholis filiformis (Roth.) C. E. Hubbard, Medicago marina L., Sporobolus virginicus (L.) Kunth, Trachomitum venetum (L.) Woodson, Polypogon maritimus Willd., Xanthium strumarium L., and Cakile maritima Scop. were abundant, as well. The average relative cover of T. venetum, which is a common perennial psammophyte, was 6.87%. The coverage of this species was determined to be 17% in the abandoned field (quadrat 4). This indicates that some of the NDV elements can regenerate when agricultural practices are abandoned.
Differences were detected in the floristic composition of the vascular plant coverage in the quadrats selected from the abandoned fields. This difference occurs with an increase of weeds and a decrease of native psammophytes in terms of species richness and coverage (%).
DCA was used to give a clear ordination of the TWINSPAN groups and sampling quadrats along the environmental variables and land-use gradients (Figure 3).
The vertical axis (axis 1) represents the distribution of 3 plant communities in relation to the land-use gradient. Undisturbed foredune plant communities with lower scores, humid dune slack, salt pioneer sward, and disturbed foredune communities with relatively higher scores are located on the vertical axis. Group A, with quadrats 4 and 5, shows the highest value of relative weed species coverage, which were classified as SV elements. The group with the higher scores on the vertical axis represents disturbed foredune communities associated with agricultural land use. Group B (quadrats 1, 2, 3, and 6) represents undisturbed and seminatural sites. Relative species coverage of SV is 39% while NDV is 36% in quadrat
1, which was identified as a seminatural site. Group C (quadrats 7 and 8) contains halophytic communities, which are classified as NDV elements, while a few SV species are associated with this group.
Quadrats 2 and 3 were located at the crossing point of 2 axes, representing undisturbed foredune communities with typical psammophytes (Euphorbia peplis L. and Medicago marina L.). Therefore, natural gradients were identified along the horizontal axis (axis 2), which represents differences in life forms and site conditions.
The DCA ordination showed that differences in the distribution of plant communities along the disturbance gradient are strongly influenced by the land use. The rates
of NDV / SV in terms of species richness (in number) and relative species cover (%) support the above-mentioned assertion. These values show differences from undisturbed sites to abandoned fields. In undisturbed sites (quadrats 2, 3, 6, and 7), the average of NDV / SV was 9 / 2, while it was 12 / 13 in the seminatural site (quadrat 1), 9 / 9 in the abandoned fields at the foredune (quadrats 4 and 5), and 6 / 4 at the salt pioneer sward (quadrat 8).
3.2. Mycorrhizal colonization
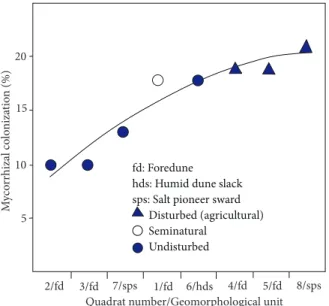
AM root colonization rates were higher in the abandoned fields than in the undisturbed dunes (Table 1), and this variation along the natural and land-use gradients is shown in Figure 4.
Figure 2. TWINSPAN groups and species classification of the plant communities. Occurrence and average relative cover
The mycorrhizal colonization rate was detected to be 22% for Xanthium strumarium L. (a common invasive weed) collected from the abandoned fields. Mycorrhizal colonization was found to be only 15% in samples of the same species collected from the undisturbed dunes. This increase is closely related to the amount of organic matter, which was determined as 0.44% in the abandoned fields and only 0.22% in the undisturbed dunes. The phosphorus (P2O5) content was 14.0 kg ha–1 in the field soils and only 10.3 kg ha–1 in the undisturbed dunes (Table 1). In quadrats 7 and 8, which were located in salt pioneer sward,
phosphorus contents showed differences (43.5 kg ha–1) when compared with the dune field soils (18.8 kg ha–1).
Relatively higher P2O5 contents were linked with an increase of mycorrhizal colonization and spore numbers in the salt pioneer sward community (quadrat 8). Mycorrhizal colonization was found to be much higher on the fibrous and rhizomatous roots than on tap and bulbous roots, respectively (Table 2). Most of the weeds have fibrous roots, which created much more suitable conditions for AM colonization than the native psammophytes. Furthermore, the highest value of AM colonization was found on the species belongs to the family Gramineae.
Figure 3. An ordination of 3 TWINSPAN groups (A, B, and C) including 8 quadrats
along the natural and land-use gradients.
In cr easin g P2 O5 & Organic ma tter 100 200 300 400 500 600 700 800 3 2 1 Foredune B 6 Humid duneslack 7 8 Salt pioneer sward Foredune A C Disturbed (agricultural) Seminatural Undisturbed Humid Hallophytes Increasing salinity Psammophytes Increasing pH Dry 300 250 200 150 100 50 Decr easi ng La nd-use gradien t Nat ural-S emina tura l Ab on do ned field Natural gradient
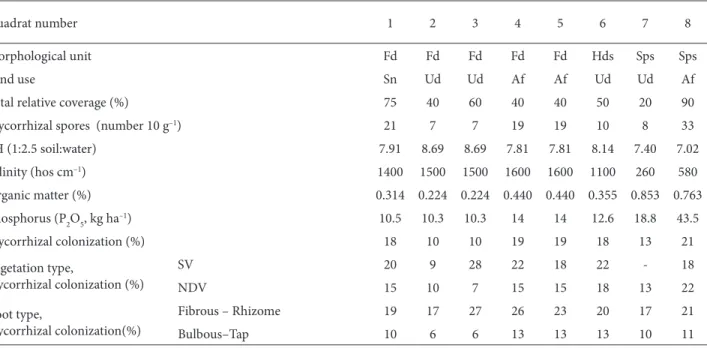
Table 1. The site conditions, measured parameters, and mycorrhizal status detected in the sampling quadrats (Fd: Foredune, Hds: humid
dune slack, Sps: salt pioneer sward, Sn: seminatural, Ud: undisturbed, Af: abandoned field).
Quadrat number 1 2 3 4 5 6 7 8
Morphological unit Fd Fd Fd Fd Fd Hds Sps Sps
Land use Sn Ud Ud Af Af Ud Ud Af
Total relative coverage (%) 75 40 60 40 40 50 20 90
Mycorrhizal spores (number 10 g–1) 21 7 7 19 19 10 8 33
pH (1:2.5 soil:water) 7.91 8.69 8.69 7.81 7.81 8.14 7.40 7.02 Salinity (hos cm–1) 1400 1500 1500 1600 1600 1100 260 580 Organic matter (%) 0.314 0.224 0.224 0.440 0.440 0.355 0.853 0.763 Phosphorus (P2O5, kg ha–1) 10.5 10.3 10.3 14 14 12.6 18.8 43.5 Mycorrhizal colonization (%) 18 10 10 19 19 18 13 21 Vegetation type, mycorrhizal colonization (%) SV 20 9 28 22 18 22 - 18 NDV 15 10 7 15 15 18 13 22 Root type, mycorrhizal colonization(%) Fibrous – Rhizome 19 17 27 26 23 20 17 21 Bulbous–Tap 10 6 6 13 13 13 10 11
3.3. Relationship between soil parameters and land use and species richness
The contents of organic matter and phosphorus were relatively higher in the abandoned fields. Salinity increases while the pH level decreases in humid conditions (Figure 3). The results showed that regeneration of the salt pioneer sward vegetation was much stronger in comparison with the foredune community due to increased salinity and humidity conditions. This can be attributed to low adaptation capacity of the invasive weeds to high salinity conditions.
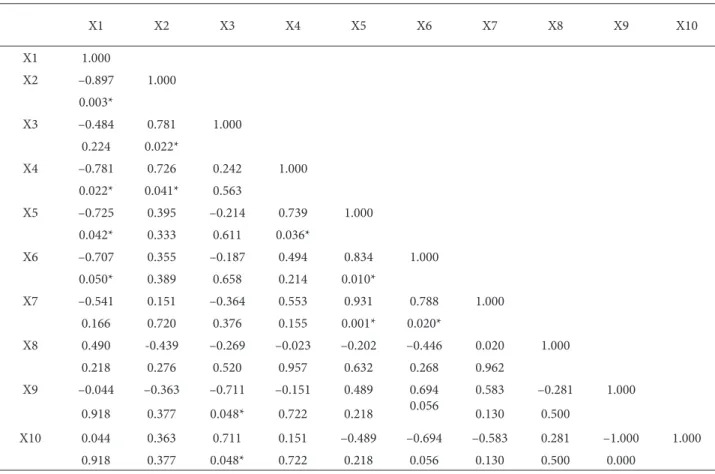
Statistically, the most significant correlation (r2 = 0.931) was observed between relative coverage of SV and number of mycorrhizal spores (X5 and X7 in Table 2). There was also a positive correlation between SV and mycorrhizal root colonization. A negative correlation was found between organic matter and soil pH.
4. Discussion
Clear changes were detected in the abundance of mycorrhizal spore and root colonization, as well as some soil properties of the dunes in the postcultivation phase. Both the floristic composition of the vascular plant communities and the rate of mycorrhizal root colonization were different between the abandoned fields and undisturbed sites. A significant correlation was found between mycorrhizal root colonization and the species richness of vascular plants. As indicated by Maremmani et al. (2003), the increase in the mycorrhizal colonization rate was associated with and positively correlated with an increase in floristic richness and plant coverage. It is obvious that vascular plant richness encourages mycorrhizal fungi species richness, as well.
In general, the number of mycorrhizal spores and species richness is higher in uncultivated soil than in cultivated soil. In a similar work, Tchabi et al. (2009) reported that species richness of mycorrhizal fungi was higher in the savanna than in the yam-field soils. In the case of the present study, the number of AM spores and root colonization in the dunes were found to be related to the agricultural land use, which results in the change of the floristic composition of vegetation (Table 1). It has been indicated that the degree of structural complexity and diversity of vegetation and the degree of dune stabilization may affect the abundance of AM spore (Cordoba et al. 2001). Erikson (2001) found a similar correlation between plant species diversity (number of plant species m–2) and colonization of mycorrhiza in seminatural grasslands. The main factor in the increase of mycorrhizal root colonization is the SV species richness, which consists of introduced annual weeds with intensive fine roots (Table 1).
Bothe (2001) indicated that the salt marsh plants in Central Europe can be strongly colonized by AM fungi and that the content of fungal spores in such saline habitats can be extremely high. Salt pioneer swards located in the study site also had higher mycorrhizal colonization than the sand dunes.
Plant species’ dependency on mycorrhiza establishment varies along a continuum from obligate to facultative to nonmycorrhizal (Barni and Siniscalco 2000). Almaca and Ortaş (2010) reported that crop rotation or plant successes have a significant impact on number of spores.
In the study, the average rate of AM colonization of roots was found to be 24%. The maximum colonization rate (46%) was observed in the roots of S. virginicus while the minimum value was only 4% in the roots of Polygonum equisetiforme Sibth. and Sm. Plant families with lower mycorrhizal root colonization than the average value (24%) are Convolvulaceae and Cyperaceae. These findings are supported by the results of Koske (1988). These families contain Cressa cretica L. (9%), Ipomoea stolonifera (Cyr.) J.F.Gmelin (10%), Scirpoides holoschoenus (L.) Sojak (12%), and C. capitatus (16%). Previous studies showed that most colonizing plants in sand dunes ecosystems are heavily infected by AM fungi (Giovannetti and Nicolson 1983; Peterson et al. 1985; Puppi and Riess 1987; Koske 1988). Although plant species belonging to families Aizoaceae, Cyperaceae, Convolvulaceae, and Commelinaceae are considered mainly nonmycotrophic, Maremmani et al. (2003) reported that Calystegia soldanella (L.) R.Br. ex Roemer & J.A.Schultes, a member of the nonmicrophytic family Convolvulaceae, had no mycorrhizal colonization in sand dunes of Tuscany, Italy. However, Vardavakis (1992) stated that the same species had a colonization rate of 9.2% in a Greek sand dune. Furthermore, mycorrhizal 3/fd 7/sps 1/fd 6/hds 4/fd 5/fd 8/sps
Quadrat number/Geomorphological unit
My co rrhizal co lo niza tion (% ) 5 10 fd: Foredune
hds: Humid dune slack sps: Salt pioneer sward Disturbed (agricultural) Seminatural Undisturbed 2/fd 15 20
Figure 4. AM colonization in 8 quadrats along the natural and
colonization on C. capitatus (Cyperaceae) (6.9%) was also reported within the same study. This might be a result of being in a heavily infected plant community and the occurrence of neighboring AM plant species.
There are a number of terrestrial, nonmycorrhizal plants, and there are several factors contributing to this condition. First, many short-lived annuals are nonmycorrhizal. The reason for this is that mycorrhizal infection typically take weeks or months to form, and the plant may finish most of its lifecycle in that time. Given that mycorrhizae require a considerable expenditure of resources before they start returning nutrients to the plant, a short lifespan indicates loss of the mycorrhiza. However, grasses and many other monocots have dense, fibrous root masses that can be either obligatorily or facultatively mycorrhizal, depending on ecotype and site conditions.
Erikson (2001) observed that the interaction among site history, nutrient status, and mycorrhiza explained the high species diversity usually found in seminatural grassland. Highly significant differences were detected in AM
colonization between the sites with different management history. Individuals from the sites with a long continuous management regime had a higher colonization rate than the individuals from sites with short or interrupted management regime. There probably is a rapid arrival of potential hosts during the first years of abandonment, and, in consequence, the propagule density is high due to the incorporation of plants, such as weed species, that stimulate the production of propagules (Jordan et al. 2000). Guadarrama et al. (2008) also concluded that traditional slash-and-burn agricultural practices (low-input agriculture) do not eliminate AM propagules and, in consequence, do not affect their dynamics, which may support the natural plant community regeneration.
Orlowska et al. (2002) reported that, although Biscutella laevigata L. is a member of Brassicaceae, which known as a nonmycorrhizal family, this species formed AM symbiosis on both heavy metal-contaminated and noncontaminated sites. Besides vesicles and coils, arbuscules were also observed, especially in roots collected prior to seed maturity.
Table 2. The correlation between site conditions, vegetation characteristics, and the data on mycorrhizal colonization.
X1 X2 X3 X4 X5 X6 X7 X8 X9 X10 X1 1.000 X2 –0.897 1.000 0.003* X3 –0.484 0.781 1.000 0.224 0.022* X4 –0.781 0.726 0.242 1.000 0.022* 0.041* 0.563 X5 –0.725 0.395 –0.214 0.739 1.000 0.042* 0.333 0.611 0.036* X6 –0.707 0.355 –0.187 0.494 0.834 1.000 0.050* 0.389 0.658 0.214 0.010* X7 –0.541 0.151 –0.364 0.553 0.931 0.788 1.000 0.166 0.720 0.376 0.155 0.001* 0.020* X8 0.490 -0.439 –0.269 –0.023 –0.202 –0.446 0.020 1.000 0.218 0.276 0.520 0.957 0.632 0.268 0.962 X9 –0.044 –0.363 –0.711 –0.151 0.489 0.694 0.583 –0.281 1.000 0.918 0.377 0.048* 0.722 0.218 0.056 0.130 0.500 X10 0.044 0.363 0.711 0.151 –0.489 –0.694 –0.583 0.281 –1.000 1.000 0.918 0.377 0.048* 0.722 0.218 0.056 0.130 0.500 0.000
X1 = pH, X2 = organic matter, X3 = salinity, X4 = P2O5, X5 = mycorrhizal spore, X6 = mycorrhizal colonization, X7 = relative cover of SV (%), X8 = relative cover of NDV (%), X9 = relative cover of annuals (%), X10 = relative cover of biannual and perennials (%), * = P < 0.05 (significance level).
It is known that Chenopodiaceae and Brassicaceae are nonmycorrhizal. Allen et al. (1989) argued that Atriplex gardneri (Moq.) D. Dietr. was vesicular-arbuscular mycorrhizal as arbuscules could be found during a short period (2–3 weeks) of the year and even the nonmycotrophic annual Salsola kali L. formed arbuscules for short intervals (2–3 days) as seedlings. Alternatively, Allen and Allen (1990) reported that limited mycorrhizal fungal activity of the perennial Atriplex confertifolia (Torr. & Frem.) J. Wats. could be found, but only when the mycorrhizal fungi were associated with a neighboring mycorrhizal grass.
Differences in soil nutrient status, particularly the amount of phosphorus between abandoned fields and undisturbed control sites, were observed. It was measured at 14 kg ha–1 in quadrats 4 and 5, while it ranged between 10.3 and 12.6 kg ha–1 in undisturbed foredune and humid dune slack areas. It is well known that the organic matter and phosphorus are higher in pioneer swards than sand dunes. The amount of phosphorus in the abandoned fields located in the salt pioneer sward was measured at 43.5 kg ha–1, while it was only 18.8 kg ha–1 in the undisturbed site. All these findings indicate that agricultural activities, including overfertilization, result in changes in the amount of organic matter and phosphorus in the dunes. It was stated that the most effective factor in the change of floristic composition of dunes is seed transportation by manure, which is used for fertilization (Yılmaz 2002). Together with the elevated amount of organic matter and phosphorus in the dunes, the introduction of seeds causes a rapid weed colonization that promotes further development of mycorrhizal symbiosis.
Isobe et al. (2008) reported that there was a significant difference in spore density among the sampling sites. In the present experiment, there was a positive correlation between extractable P and mycorrhizal spores. Since sand dune soils have low extractable P, there may be a threshold for the P level. It was reported that the density of AM fungal spores in the soybean fields was negatively correlated with the available phosphorus content (Isobe et al. 2008). The lower concentrations of soil phosphorus may stimulate
AM colonization and activity (Smith and Read 2008). On the other hand, the infection ratio was positively correlated with spore density and soil pH (Table 2).
The results of Li et al. (2007) showed that the colonization of different AM structures and the spore density varied greatly among plant species both within and between different land-use types. Similarly, Oehl et al. (2009) observed that under organic farming conditions AM communities from distinct agroecosystems differed in species composition and seasonal and succession sporulation dynamics. Ramos-Zapata et al. (2011) showed seasonal mycorrhizal colonization changes in coastal dunes of Sisal, Mexico.
Nevertheless, the results of the present study represent only 2 years of succession. In order to obtain more reliable data concerning changes in the soil nutrient status of the sand dunes and its impact on plant diversity and AM colonization, long-term monitoring is essential. The results indicate that changes occurred in vascular plant species’ richness and mycorrhizal spore abundance of coastal dune vegetation following peanut cultivation on the dunes. The change in floristic composition occurred with an increase of weeds and a decrease of native psammophytes. Ecologically, it is important to protect psammophyte flora and the sand dune itself for biodiversity and sustainability. Since AM colonization plays a significant role in the process of sand dune stabilization, and it benefits dune rehabilitation and erosion control through stimulating soil aggregation, it is strongly recommended to introduce AM-inoculated indigenous dune plants to restore degraded dunes.
Acknowledgments
The Rhizosphere Laboratory staff of the Soil Science and Plant Nutrition Department of Çukurova University is acknowledged for their assistance in root and soil analyses. We are grateful for the financial support offered by the Research Unit of Çukurova University. We also thank the anonymous reviewers for their excellent comments, and Semra Demir, the field editor.
References
Abbott LK, Robson ADJ, Asper DA, Gazey C (1992) What is the role of VA mycorrhizal hyphae in soil? In: Mycorrhizas in Ecosystems (Eds. DJ Read, DH Lewis, IJ Alexander). CAB International, Wallingford, UK, pp. 37–41.
Acosta A, Blasi C, Stanisci A (2000) Spatial connectivity and boundary patterns in coastal dune vegetation in the Circeo National Park, Central Italy. J Veg Sci 11: 149–154.
Allen MF, Allen EB (1990) Carbon source of VA mycorrhizal fungi associated with Chenopodiaceae from a semi-arid steppe. Ecology 71: 2019–2021.
Allen MF, Allen EB, Friese CF (1989) Responses of the non-mycotrophic plant Salsola kali to invasion by VA mycorrhizal fungi. New Phytol 111: 45–49.
Almaca A, Ortaş İ (2010) Growth response of maize plants (Zea
mays L.) to wheat and lentil pre-cropping and to indigenous
mycorrhiza in field soil. Span J Agric Res 8: 131–136.
Barbour MG, De Jong TM, Pavlick BM (1985) Marine beach and dune plant communities. In: Physiological Ecology of North American Plant Communities (Eds. BF Chabot, HA Mooney). Chapman and Hall, New York, London, pp. 297–322.
Barni E, Siniscalco C (2000) Vegetation dynamics and arbuscular mycorrhiza in old-field successions of the western Italian Alps. Mycorrhiza 10: 63–72.
Beena KR, Raviraja NS, Arun AB, Sridhar KR (2000) Diversity of arbuscular mycorrhizal fungi on the coastal sand dunes of the West Coast of India. Curr Sci India 79: 62–65.
Bothe H (2001) Arbuscular mycorrhiza in saline habitats. In: Book of Abstracts of the Workshop on Managing Arbuscular Mycorrhizal Fungi for Improving Soil Quality and Plant Health in Agriculture, Adana, pp.36.
Çakan H, Karataş C (2006) Interactions between mycorrhizal colonization and plant life forms along the successional gradient of coastal sand dunes in the eastern Mediterranean, Turkey. Ecol Res 21: 301–310.
Çakan H, Yilmaz KT, Düzenli A (2005) First comprehensive assessment of the conservation status of the flora of the Çukurova Deltas, Southern Turkey. Oryx 39: 17–21.
Clough KS, Sutton JC (1978) Direct observation of fungal aggregates in sand dune soil. Can J Microbiol 24: 333–335.
Cordoba AS, de Mendonca MM, Stürner SL, Rygiewicz PT (2001) Diversity of arbuscular mycorrhizal fungi along a sand dune stabilization gradient: a case study at Praia da Joaquina, Ilha de Santa Catarina, South Brazil. Mycoscience 42: 379–387. Corkidi L, Rincon E (1997) Arbuscular mycorrhiza in a tropical sand
dune ecosystem on the Gulf of Mexico, I. Mycorrhizal status and mycorrhizal inoculum potential along a successional gradient. Mycorrhiza 7: 9–15.
Davis PH (1965–1988) Flora of Turkey and the East Aegean Islands. Edinburgh University Press, Edinburgh.
Erikson A (2001) Arbuscular mycorrhiza in relation to management history, soil nutrients and plant species diversity. Plant Ecol 155: 129–137.
Forster SM (1979) Microbial aggregation of sand in a maritime sand dune succession. Soil Biol Biochem 13: 537–543.
Forster SM, Nicolson TH (1981a) Microbial aggregation of sand in a maritime sand dune succession. Soil Biol Biochem 13: 205– 208.
Forster SM, Nicolson TH (1981b) Aggregation of sand from a maritime embryo sand dune by microorganisms and higher plants. Sol Biol Biochem 13: 199–203.
Gerdemann JW, Nicolson TH (1963) Spores of mycorrhizal endogamy species extracted from soil by wet sieving and decanting. Trans Brit Mycol Soc 46: 235–244.
Giovannetti M, Mosse B (1980) An evaluation of techniques for measuring vesicular arbuscular mycorrhizal colonization in roots. New Phytol 84: 489–500.
Giovannetti M, Nicolson TH (1983) Vesicular-arbuscular mycorrhizas in Italian sand dunes. Trans Br Mycol Soc 80: 552–557.
Guadarrama P, Castillo-Argüero S, Ramos-Zapata JA, Camargo-Ricalde SL, Álvarez-Sánchez J (2008) Propagules of arbuscular mycorrhizal fungi in a secondary dry forest of Oaxaca, Mexico. Rev Biol Trop 56: 269–277.
Hill MO (1974) Correspondence analysis: a neglected multivariate method. JR Statist Soc Ser C 23: 340–354.
Isobe K, Sugimura H, Maeshima T, Ishii R (2008) Distribution of arbuscular mycorrhizal fungi in upland field soil of Japan - 2. Spore density of arbuscular mycorrhizal fungi and infection ratio in soybean and maize fields. Plant Prod Sci 11: 171–177. Jordan NR, Zhang J, Huerd S (2000) AM fungi potential roles in
weed management. Weed Res 40: 397–410.
Koske RE (1975) Endogone spores in Australian sand dunes. Can J Bot 53: 668–672.
Koske RE (1988) Vesicular-arbuscular mycorrhiza of some Hawaiian dune plants. Pac Sci 42: 217–229.
Koske RE, Gemma JN (1989) A modified procedure for staining roots to detect VAM. Mycol Res 92: 486–505.
Koske RE, Gemma JN (1995) Vesicular-arbuscular mycorrhizal inoculation of Hawaiian plants: a conservation technique for endangered tropical species. Pac Sci 49:181–191.
Koske RE, Polson WR (1984) Are VA mycorrhiza required for sand dune stabilization? Bioscience 34: 420–424.
Li LF, Zhang Y, Zhao ZW (2007) Arbuscular mycorrhizal colonization and spore density across different land-use types in a hot and arid ecosystem, Southwest China. J Plant Nutr Soil Sc 170: 419–425.
Little LR, Maun MA (1996) The “Ammophila problem” revisited: a role for mycorrhizal fungi. J Ecol 84: 1–7.
Maremmani A, Bedini S, Matosevic I, Tomei PE, Giovannetti M (2003) Type of mycorrhizal associations in two coastal nature reserves of the Mediterranean basin. Mycorrhiza 13: 33–40. Maun M A (1994) Adaptations enhancing survival and establishment
of seedlings on coastal dune systems. Plant Ecol 111: 50–70. McLean EO (1982) Chemical and microbiological properties. In:
Methods of Soil Analysis, 2nd ed. (Eds. LA Page, RR Miller, DR Keeney). American Society of Agronomy. Madison, Wisconsin, USA, pp. 199–224.
Murphy Y, Riley JPA (1962) Modified single solution method for determination of phosphate in natural waters. Anal Chem Acta 2: 31–36.
Oehl F, Sieverding E, Ineichen K, Mader P, Wiemken A, Boller T (2009) Distinct sporulation dynamics of arbuscular mycorrhizal fungal communities from different agroecosystems in long-term microcosms. Agr Ecosyst Environ 134: 257–268. Orlowska E, Zubek S, Jurkiewicz A, Szarek-Lukaszewska G, Turnau
K (2002) Influence of restoration on arbuscular mycorrhiza of
Biscutella laevigata L. (Brassicaceae) and Plantago lanceolata L.
(Plantaginaceae) from calamine spoil mounds. Mycorrhiza 12: 153–160.
Ortaş I (2002) Soil biological degradation. In: Encyclopedia of Soil Science (Ed. R. Lal). Marcel Dekker, New York, pp. 264–267. Ortas I, Harris PJ, Rowell DR (1996) Enhanced uptake of phosphorus
by mycorrhizal sorghum plants as influenced by forms of nitrogen. Plant Soil 184: 255–264.
Peterson RL, Ashford A, Allway G (1985) Vesicular-arbuscular mycorrhizal associations of vascular plants on Heron Island, a greater barrier coral cay. Aust J Bot 33: 669–676.
Puppi G, Riess S (1987) Role and ecology of VA mycorrhiza in sand dunes. Angew Bot 61:115–126.
Ramos-Zapata JA, Zapata-Trujillo R, Ortiz-Diaz JJ, Guadarrama P (2011) Arbuscular mycorrhizas in a tropical coastal dune system in Yucatan, Mexico. Fungal Ecol 4: 256–261.
Read DJ (1989) Mycorrhizas and nutrient cycling in sand dune ecosystem. In: Proceedings of the Royal Society of Edinburgh (Eds. CH Gimingham, W Ritchie, BB Willetts, AJ Willis), Vol. 96, Edinburgh, pp. 89–100.
SAS/STAT Institute (1987) Guide for Personal Computers. Version 6. SAS, Inc., Cary, North Carolina, USA.
Smith SE, Read DJ (2008) Mycorrhizal Symbiosis. Academic Press, London.
Sutton JC, Sheppard BR (1976) Aggregation of sand dune soil by endomycorrhizal fungi. Can J Bot 54: 326–333.
Sylvia DM (1989) Nursery inoculation of sea oats with vesicular-arbuscular mycorrhizal fungi and out planting performance on Florida beaches. J Coastal Res 5: 747–754.
Sylvia DM, Jarstfer AG (1997) Distribution of mycorrhiza on competing pines and weeds in a southern pine plantation. Soil Soc Am J 61: 139–144.
Tchabi A, Burger S, Coyne D, Hountondji F, Lawouin L, Wiemken A, Oehl F (2009) Promiscuous arbuscular mycorrhizal symbiosis of yam (Dioscorea spp.), a key staple crop in West Africa. Mycorrhiza 19: 375–392.
van der Koornhuyse P, Ridgway KP, Watson IJ, Filter AH, Young JPW (2003) Co-existing grass species have distinctive arbuscular mycorrhizal communities. Mol Ecol 12: 3085–3095.
van der Meulen F, Salman HPM (1996) Management of Mediterranean coastal dunes. Ocean Coast Manage 30: 177–195.
Vardavakis E (1992) Mycorrhizal endogonaceae and their seasonal variations in a Greek sand dune. Pedobiologia 36: 373–382. Yılmaz KT (2002) Evaluation of the phytosociological data as a tool
for indicating coastal dune degradation. Isr J Plant Sci 50: 229–238.
Yılmaz KT, Alphan H, İzcankurtaran Y (2004) Forest stand structure along an altitudinal gradient in the ICCAP area. In: Proceedings of the International Workshop for the Research Project on the Impact of Climate Change on Agricultural Production System in Arid Areas (ICCAP) (Eds. T Watanabe, R Kanber), pp. 112–117.