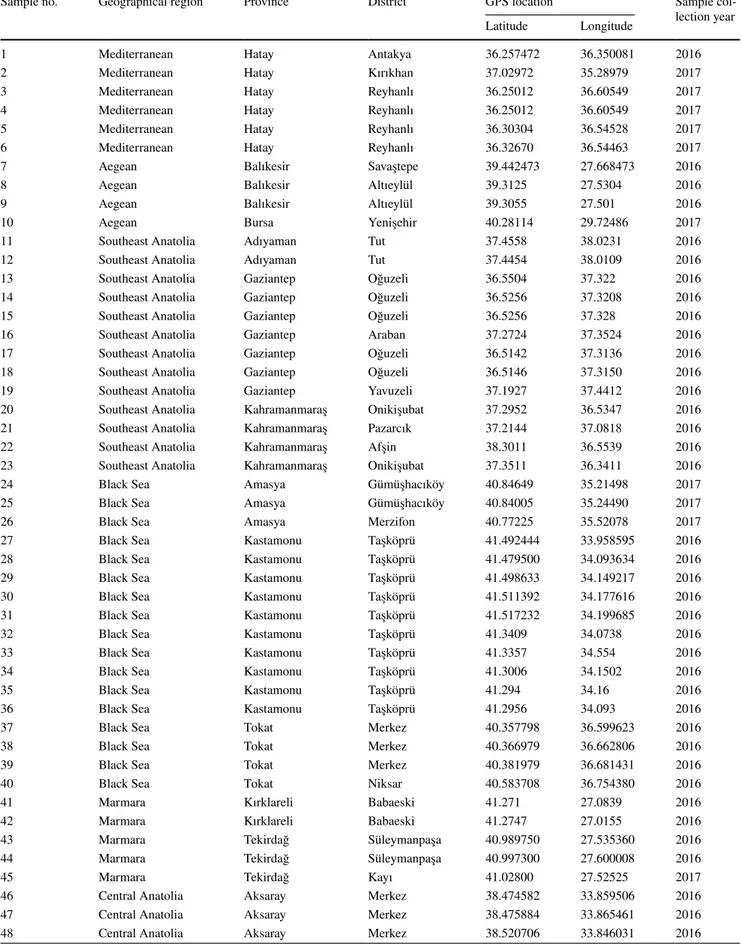
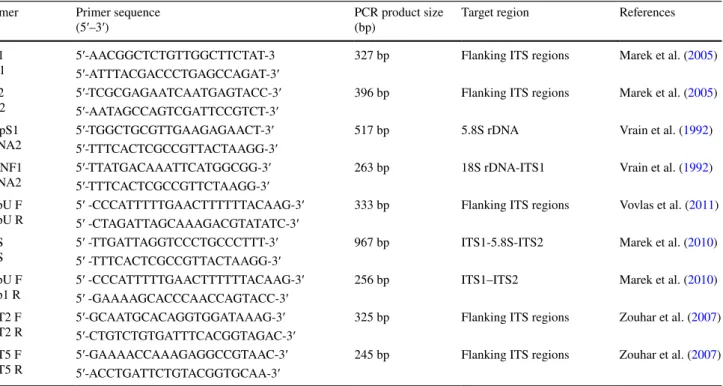
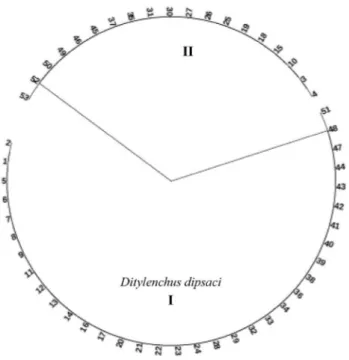
Molecular characterization of Ditylenchus dipsaci on garlic in Turkey
Tam metin
Şekil
Benzer Belgeler
Our study aimed to test the susceptibilities of 50 Candida species isolated from blood cultures against anidulafungin, micafungin, caspofungin, flucytosine,
Çalışmada işletme verimliliğinin ölçülmesinde kullanılan parametrik olmayan matematiksel programlama tabanlı etkinlik ölçüm yöntemlerinden Veri Zarflama Analizi
For the case with a known receiver height and a uniform circular layout for the PDs (which is a common and efficient configuration, as investigated in [28]), a compact CRLB
Programda ne hedef ne de davranış özelliği gösteren öğrenci kazanımlarının birçoğu ise birden fazla yüklem aynı kazanım tümcesinde yer aldığı için
Bu modellerden hangisinin doğru olduğunu görebilmek için Plüton’un Güneş’ten uzaklaşırken incelenmesinin gerekli olduğunu söyleyen gökbilimciler, NASA’nın New
vulnus obtained from wheat and barley cultivated fields in 2016 to 2017 in Turkey (Karaman and Konya provinces) were identified using real-time PCR and melting curve
Genel olarak içerisinde % 10’dan daha az CaO içerdikleri için düşük kireçli uçucu kül olarak isimlendirilir.. Kendi kendine sertleşme özelliğine
KSS kavramının bir şirketin kime karşı sorumlu olduğunu açıkça cevaplayamadığından yakınan Carroll (1991: 43), ST’nin; işletme için toplumdaki en zorunlu
