Laboratuvar koşullarında yüksek glukoza maruz kalmış insan eritrositlerinde kondroitin-4-sülfatın antioksidan savunma sistemi üzerine etkisi
Tam metin
Şekil

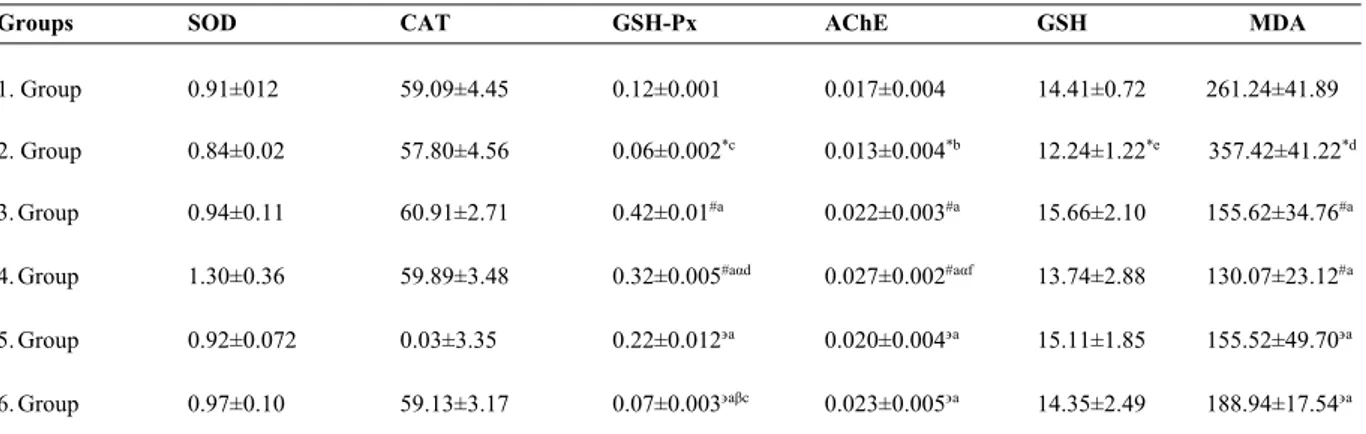
Benzer Belgeler
Görünüş : Aşırı aşınma ve merkezleme pimi karşısındaki çalışma yüzeyi üzerinde tutukluk başlangıcı ve yatak üzerinde pim izleri. Merkezleme pimi çok uzun
In this study, we investigated the effects of pulsatile and nonpulsatile perfusion on hemolysis parameters, protection of blood components, transfusion requirement,
With flooding, it is possible to reach a destination node from a source node with minimum number of hops, with minimum delay but with maximum network traffic.. Figure 1
■陸生來臺輔導說明會 本校今年首度招收 7 位大學部陸生(3 女、4 位男),分別來自上海、廣東、江
Ethanolic extract of adlay bran and residue from ethanolic extraction of adlay bran significantly suppressed the formation of ACF in the distal colons but did not aff ect the
2014, non-pregnant pa- tients over the age of 18 who were not diagnosed with diabetes and prediabetes who applied Health Sciences University Sisli Hamidiye Etfal Training and
Assuming the heat transfer from the surface to be uniform, determine, a.(05) the amount of heat the iron dissipates during a 2-hour period, in kWh, b.(05) the heat flux on the
Our findings demonstrated that although the absolute values of the 2DSTE-derived indices of the longitudinal deformation of the LV myocardium (i.e., systolic strain, strain rate,
